
Gastrophysa janthina (Suffrian, 1851)
|
publication ID |
https://doi.org/10.5281/zenodo.178650 |
|
DOI |
https://doi.org/10.5281/zenodo.6244699 |
|
persistent identifier |
https://treatment.plazi.org/id/D02CFF38-FF85-C71D-91B5-535EFB9F1D2A |
|
treatment provided by |
Plazi |
|
scientific name |
Gastrophysa janthina (Suffrian, 1851) |
| status |
|
Gastrophysa janthina (Suffrian, 1851) , mature larva
( Figs. 1–6 View FIGURE 1 View FIGURE 2 – 7 , 8 View FIGURE 8 )
Material examined. SPAIN. A Coruña, A Capela, Caaveiro, (UTM 29TNJ7407), 50 m asl, 11 April 1999, 5 mature larvae; Ourense, Chandrexa de Queixa (UTM 29TPG3478), 600 m asl, 8 May 1999, 14 mature larvae. Voucher specimens are deposited in the Museo Nacional de Ciencias Naturales, Madrid, Spain.
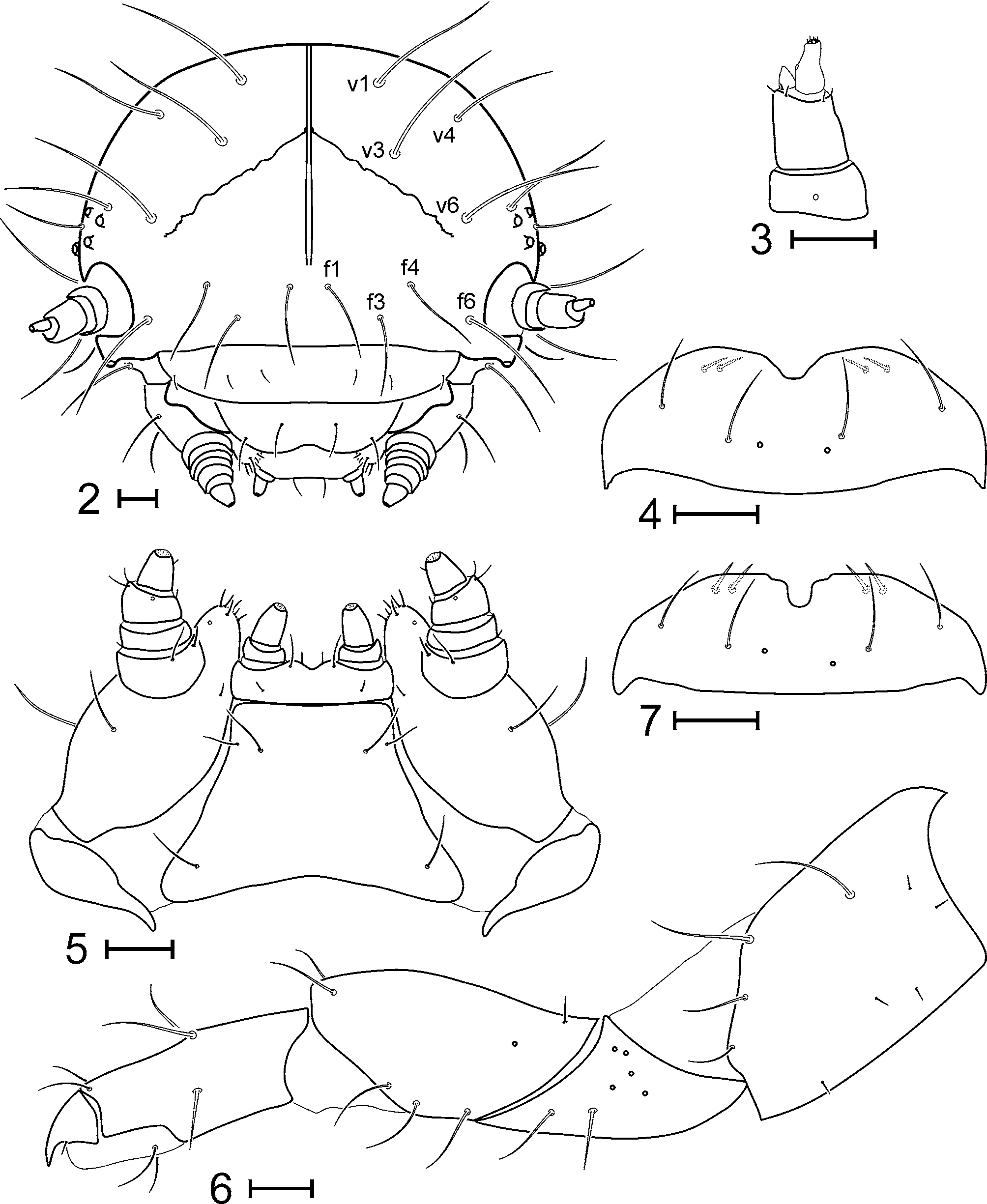
Description. Habitus as in Fig. 1 View FIGURE 1 . Length: 5.5–7.1 mm. Body eruciform, moderately convex and slightly arched in preserved specimens. Inter-tubercular plates and tubercles concolor, brown in dorsal region, light brown in ventral region. Head. Hypognathous, well sclerotized. Color brown. Epicranial suture well developed and long, frontal arms distinct, V-shaped and almost straight ( Fig. 2 View FIGURE 2 – 7 ). Endocarina present, extending to the middle of the frons and thus not reaching the clypeus. Vertex bearing 4 pairs of setae ( v1, v3, v4, v6).
Frons with 4 pairs of setae (f1, f3, f4, f6). Antennae short and well sclerotized, three-segmented ( Fig. 3 View FIGURE 2 – 7 ): first joint highly transverse, bearing 2 placoid sensilla (only 1 visible in frontal view); second joint 1.5 times longer than wide, bearing a large conical membranous sensillum and 5 minute setae; distal joint narrow, bottleshaped, constricted anteapically, with membranous apex bearing 6 highly minute setae similar in size (no central one larger than the others) and 1 placoid sensillum slightly produced and situated before the anteapical constriction. Stemmata arranged in two groups, 1 pair located below the base of antenna and 2 pairs behind the antenna. Clypeus with 2 pairs of setae. Mouthparts. Labrum ( Fig. 4 View FIGURE 2 – 7 ) bearing 1 pair of placoid sensilla and 2 pairs of setae on dorsal surface, one near middle, the other near lateral sides; anterior border with a median wide V-shaped notch; ventral surface with 2 pairs of short setae not reaching the anterior border. Mandibles symmetrical, 5-toothed, bearing 2 setae on external face and 1 placoid sensillum on dorsal side. Maxillae ( Fig. 5 View FIGURE 2 – 7 ): cardo transverse, without setae; stipes elongate, with 2 large setae on external face and 1 short seta on the internal one; mala bearing 10 setae on internal margin and apex, basal setae longer than apical ones; maxillary palpi 4-segmented, first joint almost as long as wide, bearing 2 long setae basally in ventral side, second joint transverse with 1 minute seta on external side, third joint longer than second, but still transverse, with 1 seta on internal face, 1 placoid sensillum on ventral face and two setae on external side, and fourth joint conical with 1 minute seta on internal face and membranous apex bearing 9–11 highly minute setae. Labium ( Fig. 5 View FIGURE 2 – 7 ) with postmentum membranous, bearing 3 pairs of setae, anterolateral one shorter than the others; prementum with 4 pairs, 2 posterior and 2 anterior to labial palpi (only one visible in Fig. 5 View FIGURE 2 – 7 ), one of the former much longer than the others; labial palpi 2-segmented, first joint transverse, distal joint conical with membranous apex bearing 9 highly minute setae. Thorax. Prothorax with tubercles D (dorsal), DL (dorsolateral) and EPa (epipleural) fused together in a pronotal sclerite ( Fig. 8 View FIGURE 8 ), pronotum (D-DL-EPa) bearing 10 pairs of primary setae along with 1 pair of extremely minute ones; tubercle EPp bearing 1 setae; tubercle P (pleural) with 1 setae; ventral region with midventral tubercle ES (eusternal) fused with tubercle SS (sternellar), tubercle SS- ES bearing 2 pairs of setae. Meso- and metathorax with 5 tubercles on each side of dorsal region: Dai (dorsal anterior interior, bearing 1 seta), Dae (dorsal anterior exterior, 1 seta), Dpi-Dpe (dorsal posterior interior and dorsal posterior exterior fused together, 1 seta), DLai-DLpi (dorsolateral anterior interior and dorsolateral posterior interior fused together, 1 long seta and 2 smaller ones), and DLae-DLpe (dorsolateral anterior exterior and dorsolateral posterior exterior fused together, 2 large setae along with 1–2 minute ones) enclosing the eversible gland; epipleural region with 2 tubercles, EPa (epipleural anterior, 2–3 setae, variable among individuals and even lateral sides of a specimen) and EPp (epipleural posterior, 1 seta); mesothoracic spiracle (st) very close to EPa tubercle; P tubercle bearing 1 seta; SS-ES bearing 2 pairs of setae. Legs. All pairs similar in size; trochantin ( Tr) located in front of P tubercle ( Fig. 8 View FIGURE 8 ), bearing 2 extremely minute setae in anterior half; prothoracic trochantin also bearing a large seta near posterior angle; coxa 1.5 times longer than wide in lateral view ( Fig. 6 View FIGURE 2 – 7 ), with 4 large setae on dorsal half of anterior side and 5 shorter ones in lateral declivities; trochanter triangular in lateral view, with 2 setae on each side, 5 placoid sensilla near coxal articulation on anterior side and 2 on posterior side; femur trapezoidal in lateral view, with 3 setae dorsally (1 pair near apex and 1 single minute one near base, 3 ones on each lateral declivity and 1 placoid sensillum on anterior side; tibiotarsus twice longer than wide, bearing 4 setae dorsally, 1 on each side and 2 ventrally; unguis wide basally, curved apically, bearing 1 seta on lower side. Abdomen. Segments 1–6 with only 3 tubercles on each side of dorsal region ( Fig. 8 View FIGURE 8 ): Dpi-Dpe (2 setae), Dae (1 seta) and DLae-DLpe (2 setae) enclosing the eversible gland; Dai and DLai-DLpi lacking; epipleural region with tubercle EP bearing 2 setae; spiracle (st) located between EP and DLae-DLpe tubercles, closer to EP; P tubercle with 2 setae, one of them shorter (lacking in segment 1); sternal region presents the following tubercles: PS (parasternal, 2 setae, excepting segment 1 with only 1 seta), SS (1 seta), ES (1 pair of setae); secondary tubercle as1 (with 1 minute seta) present. Segment 7 with tubercles Dpi-Dpe fused together in a single unpaired dorsal sclerite, and with tubercles as1, PS, SS and ES fused together in a ventral sclerite; segment 8 and 9 with dorsal and dorsolateral tubercles fused together, as well as pleural and ventral ones; segment 9 with tubercle EP lacking; segment 10 forming anal pseudopod, without dorsal tubercles, ventral ones fused.
Distribution and ecology. Although G. janthina was cited by Oliveira (1893) from “ tout le Portugal ” and after Jolivet (1951) this species is distributed throughout the Iberian peninsula, considering the available data the taxon is restricted to the northwest quadrant of the Iberian peninsula: it is very common in Galicia ( Baselga & Novoa, 2000a; 2000b; 2002; 2004a; 2006) and it was also recorded from León ( Petitpierre & Gómez-Zurita, 1998), North Portugal ( Baselga & Novoa, 2000b), Zamora ( Baselga & Novoa, 2003), Asturias and Cantabria ( Petitpierre, 2005). In all these citations the species is referred as G. unicolor . Gastrophysa janthina lives on Polygonaceae of the genera Polygonum L. (Petitpierre & G³mez-Zurita, 1998) and Rumex L. ( Baselga & Novoa, 2000a; 2000b; 2002; 2003), but no host plant was ever identified to the specific level. The larvae described in this paper were collected on Rumex spp. (field identification).
Discussion. Four species of Gastrophysa are distributed in the western Palaearctic region ( Warchalowski, 2003) but the larval stages are known only for G. polygoni and G. v i r i d u l a ( Hennig, 1938; Steinhausen, 1994). The larvae of the genus Gastrophysa are well defined by the following combination of characters ( Cox, 1982; Steinhausen, 1994): body dorsally not strongly convex, epicraneal suture long, labrum with an anterior median notch and 2 pairs of setae on upper surface, pronotum bearing a low number of setae on disc, dorsolateral tubercles of meso-, metathorax and abdominal segments 1–7 with eversible glands, abdominal segments 1–6 with a dorsal anterior tubercle, tarsal claws not toothed on lower side. The mature larva of G. janthina agrees all these diagnostic characters, as described in this paper, and keys out with Gastrophysa viridula following both Hennig (1938) and Steinhausen (1994) works, due to its brown dorsum with tubercles clearly delimited from intertubercular tegument. No characters are available in the cited papers to separate G. janthina from G. viridula , but larvae of the latter species were collected [ SPAIN: Lleida, Valle de Ar<n, Circo de Colomers (31TCH2922), 23 July 2006, 19 mature larvae and 5 adults], and the comparison of the larvae has showed that they are almost identical in both species. No differences were observed in the number and configuration of tubercles or setae and even the same variation in the number of setae of meso- and metathoracical tubercles DLae-DLpe and EPa is observed in both species. The only diagnostic character that could be detected is the outline of the labrum which has a wide V-shaped median notch in G. janthina , whereas it has a narrow U-shaped notch in G. viridula ( Fig. 7 View FIGURE 2 – 7 ).
The description of the larva of G. janthina is the first one within the subgenus Exiguipenna Jolivet , which includes only this species and G. analis from the Balkan peninsula. The subgenus was established based on the reduction of wings in adults of both species. No other characters support the splitting of the genus, and the larval morphology does not supply any other diagnostic feature, providing that the two species are virtually identical at this stage, except for the shape of the labrum, which is highly variable among close species of Chrysomelinae ( Steinhausen, 1994; Baselga & Novoa, 2004b) and thus not appropriate for supporting supraspecific taxa. On the other hand, wing reduction or loss is a common evolutionary event frequently produced in independent lineages due to environmental conditions (see Wagner & Liebherr, 1992 for a review of the causes leading to apterism and its frequent independence of phylogeny; Philips, 2000 for a report of the independent origin of flightlessness in New World Ptininae ; and Flowers, 2004 for an example in other Chrysomelidae ). In my opinion it is questionable that brachypterism justify the subgenus Exiguipenna , in absence of any other evidence, and further phylogenetic research should clarify its validity.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Chrysomelinae |
|
Genus |