
Pseudaegina rhodina ( Haeckel, 1879 )
|
publication ID |
https://doi.org/ 10.35929/RSZ.0049 |
|
DOI |
https://doi.org/10.5281/zenodo.5710643 |
|
persistent identifier |
https://treatment.plazi.org/id/D0118A7C-5B5F-0042-FC52-F8C9FD327ED8 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudaegina rhodina ( Haeckel, 1879 ) |
| status |
|
Pseudaegina rhodina ( Haeckel, 1879) View in CoL View at ENA
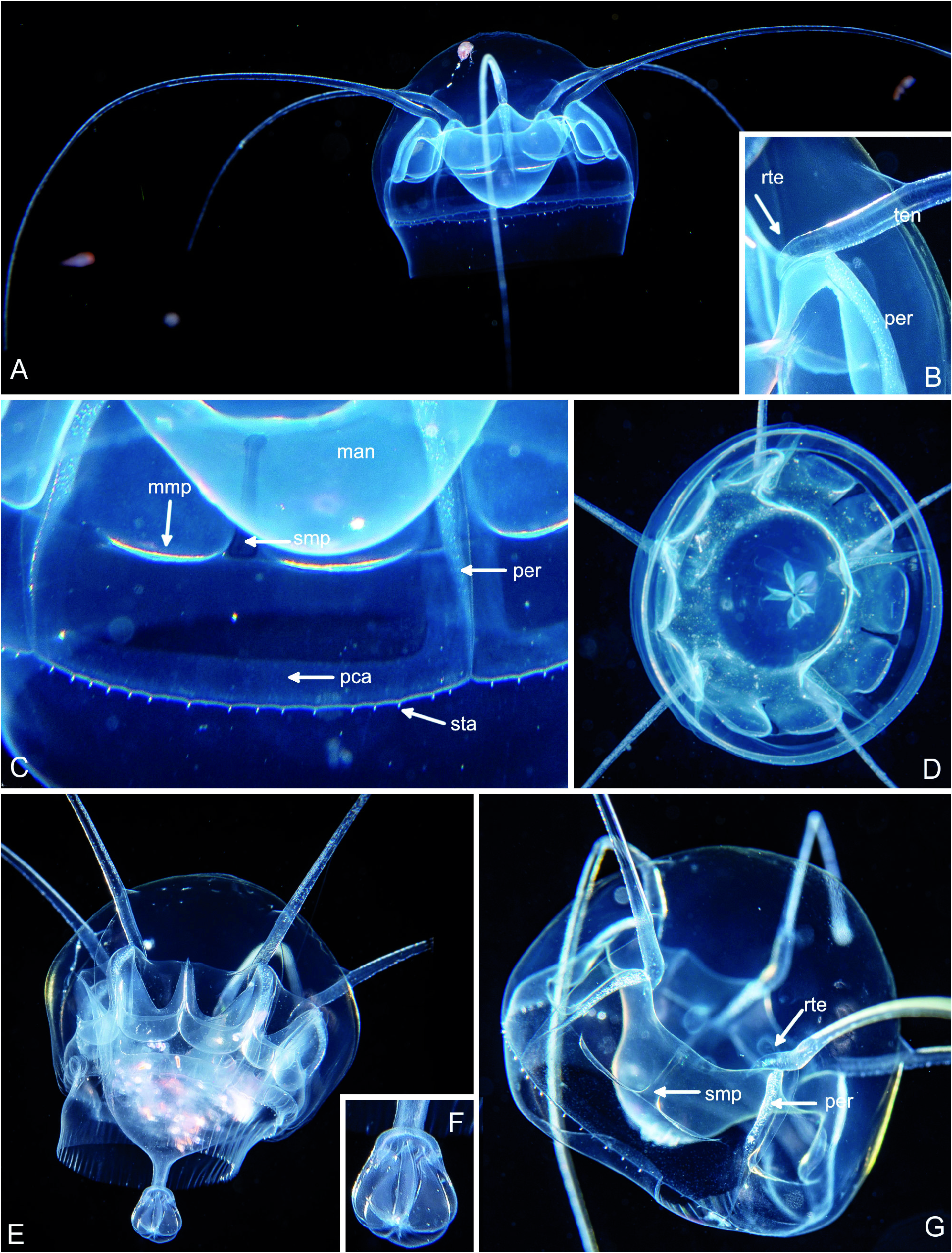
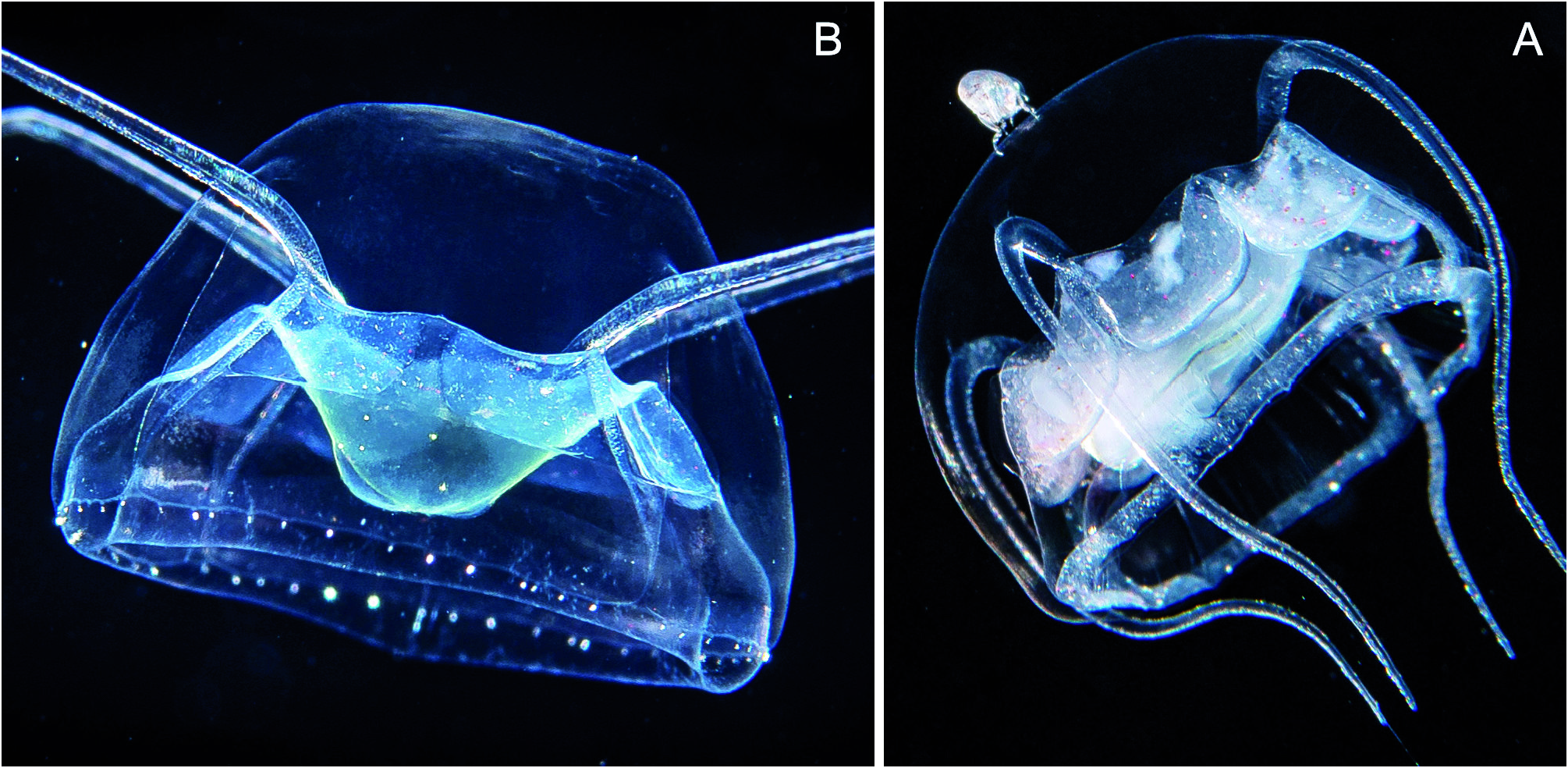
Fig 51 View Fig A-G, Fig. 52 View Fig A-B
Aegina rhodina Haeckel, 1879: 338 , pl. 20 figs 11-15. – Mayer, 1904: 27, pl. 4 figs 28-29. – Maas, 1905: 71, is Atlantic species. – Mayer, 1910: 452, plate 52, fig. 5; plate 54, figs 11, 11’-11’”.
Aegina canariensis Haeckel, 1879: 339 . – Maas, 1905: 71, synonym of A. rhodina . – Mayer, 1910: 452, synonym.
Aegina eschscholtzi Haeckel, 1879: 339 . – Maas, 1905: 71, synonym of A. rhodina . – Mayer, 1910: 453.
Solmundus tetralinus Haeckel, 1879: 351 . – Maas, 1905: 71, synonym of A. rhodina . – Mayer, 1910: 452, synonym of A. rhodina .
? Aegina lactea Vanhöffen, 1908: 50 View in CoL , pl. 1 fig. 3.
Aegina citrea View in CoL . – Bleeker & Van der Spoel, 1988: 244, figs 43- 44. – Larson et al., 1989: 789. – in part Kramp, 1959a: 61. [not Aegina citrea Eschscholtz, 1829 View in CoL ]
Pseudaegina rhodina View in CoL . ‒ Lindsay et al., 2017: 507, figs 11-13, redescription, new combination.
Examined material
Specimens with 5 tentacles: BFLA4015 ; 1 specimen; 17- JAN-2019; size 18 mm; part preserved in formalin and deposited as UF-013441 , small part in alcohol for DNA extraction; 16S sequence MW528658 View Materials . – BFLA4047 ; 1 specimen; 01-APR-2019; size 16 mm; part preserved in formalin and deposited as UF-013451 , small part in alcohol for DNA extraction; 16S sequence MW528659 View Materials . – BFLA4067 ; 1 specimen; 10-APR-2019; size 20 mm; part preserved in formalin and deposited as UF-013778 , small part in alcohol for DNA extraction; 16S sequence identical to MW528659 View Materials . – BFLA4322 ; 1 specimen; 24- JAN-2020; size 15 mm; part preserved in formalin and deposited as UF-013887 , small part in alcohol for DNA extraction; 16S sequence MW528707 View Materials . – 13-MAR-2020; 1 specimen; 15 mm; not collected.
Specimens with 4 tentacles: BFLA4120 ; 1 specimen; 04- JUN-2019; size 9 mm; part preserved in formalin and deposited as UF-013803 , small part in alcohol for DNA extraction; 16S sequence MW528678 View Materials . – BFLA4134 ; 1 specimen; 15-JUN-2019; size 11 mm; part preserved in formalin and deposited as UF-013809 , small part in alcohol for DNA extraction; 16S sequence not determinable. – BFLA4118 ; 1 specimen; 04-JUN-2019; size 9 mm; part preserved in formalin and deposited as UF-013809 , small part in alcohol for DNA extraction, not examined.
Observations
Form with 5 tentacles: Umbrella approximately hemispherical, 15-20 mm, jelly firm, apical mesoglea half the bell height, tentacles issue at about 2/3 of bell height, above tentacles a small bulge ( Fig. 51 View Fig A- B). Velum broad ( Fig. 51A View Fig ). Stomach about half the bell diameter, mounted on a rounded mesogleal cone of the same diameter reaching to level of bell margin ( Fig. 51A View Fig ). Mouth rim can form temporary folds or even a tubular extension with terminal folds ( Fig. 51 View Fig D-F). Five subdivided manubrial pockets ( Fig. 51 View Fig C-D). Manubrial pockets broadly U-shaped, reaching from stomach rim to about half the distance to bell margin ( Fig. 51A View Fig ), in middle a narrow septum which does not reach to the stomach level or level of tentacle origin ( Fig. 51C View Fig , smp). At bottom of left and right side of the pairs of pouches originates a broad peripheral canal, directed first along peronium towards the bell margin and then following the latter ( Fig. 51C View Fig ). Five tentacles originating in upper half of medusa at about upper level of stomach, the proximal end embedded in an exumbrellar furrow, the tentacle roots rather short, entering the mesoglea, tapering, and curved towards oral ( Fig. 51B View Fig ); tentacle length about 2-3 bell diameters, curved towards oral, nematocysts concentrated on upper side but also present on underside. Below tentacles a deep cleft in exumbrella reaching down to bell margin, its floor with a thickened epidermis containing nematocysts (= peronium, Fig. 51 View Fig C-G). 48 to ca. 68 statocysts. No otoporpae. Almost colourless, greenish or yellowish cast in daylight.
Form with 4 tentacles: As above, but only four tentacles and four pairs of manubrial pouches, size 9 to 11 mm, 36 to 60 statocysts.
16S data: In the maximum likelihood tree ( Fig. 48 View Fig ), all five samples clustered in a lineage that was well separated from most other Narcomedusae . The clade had a distinct internal dichotomy with the single 4-tentacled specimen being rather apart from the 5-tentacled medusae (7.6 to 7.8% base pair differences). The three haplotypes of the 5-tentacled medusae were rather similar (0.18-0.36% divergences, Table 1 View Table 1. 16 ).
Distribution: Tropical to subtropical Atlantic Ocean ( Lindsay et al., 2017). Type locality: Canary Islands, Lanzarote.
Remarks: Haeckel’s type material of this species consisted in three specimens. Two of them had four tentacles and a size of 40 mm, one had five tentacles and measured 50 mm. He depicted a mature female with four tentacles. The statocyst numbers were given as two per manubrial hemi-pouch (16-20 in total).
Kramp (1955b, 1961) synonymized this species with the similar Aegina citrea Eschscholtz, 1829 , an opinion already pondered by Mayer (1910). This view was upheld until Lindsay et al. (2017) revised Aegina citrea and split it into several species and belonging to separate families, this based on morphological and genetic differences. The main morphological difference to Aegina is the shape of the tentacle root. In Aegina they are curved towards aboral, in Pseudaegina they are curved towards oral.
Our specimens with 5 tentacles matched the description of Lindsay et al. (2017) very well as they came from the same region. There are, however, some differences to Haeckel’s description. Our specimens were smaller (up to 20 mm) but all immature. The most obvious difference is the number of statocysts: Haeckel saw about 16-20, while we found regularly 60-68.
The haplotype divergence in our sample was high (p-value up to 7.8%) and in the maximum likelihood tree the samples split into two sub-clades ( Fig. 48 View Fig ). This separation coincided with two morphotypes: the four- and five-tentacled form (see above). Unfortunately, we could get only one sequence of the four-tentacled form and due to the low number of samples, this distribution could also be due to mere contingency. The four-tentacled medusae were all smaller than those with five tentacles and it is nevertheless the possible that our material was composed of two sister-species. Because the original material of Haeckel included both four- and five-tentacled forms, it is not possible to use the tentacle number in the diagnosis of a new species without the designation of a lectotype (or a neotype as Haeckel’s material is likely lost). A population-genetic study at the type locality combined with the designation of a neotype is thus needed to resolve this taxonomic problem. Because of these unresolved issues and because our samples were monophyletic, we felt it premature to describe one of the clades as a new species.
One particularity not mentioned in previous descriptions concerns the temporary folds of the mouth region ( Fig. 51 View Fig D-F). One of the figures in Lindsay et al. (2017: fig. 12C) also shows this ruffling of the mouth margin. In one of our animals the mouth was even drawn out to a tube ending in a flower-like, folded mouth region. The folds disappeared after fixation. It is thus unlikely that they can ever be seen in standard net-plankton samples.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pseudaegina rhodina ( Haeckel, 1879 )
| Schuchert, Peter & Collins, Richard 2021 |
Pseudaegina rhodina
| Lindsay D. J. & Grossmann M. M. & Bentlage B. & Collins A. G. & Minemizu R. & Hopcroft R. R. & Miyake H. & Hidaka-Umetsu M. & Nishikawa J. 2017: 507 |
Aegina citrea
| Larson R. J. & Mills C. E. & Harbison G. R. 1989: 789 |
| Van Der Spoel S. & Bleeker J. 1988: 244 |
| Kramp P. L. 1959: 61 |
Aegina rhodina
| Mayer A. G. 1910: 452 |
| Maas O. 1905: 71 |
| Mayer A. G. 1904: 27 |
| Haeckel E. 1879: 338 |
Aegina canariensis
| Mayer A. G. 1910: 452 |
| Maas O. 1905: 71 |
| Haeckel E. 1879: 339 |
Aegina eschscholtzi
| Mayer A. G. 1910: 453 |
| Maas O. 1905: 71 |
| Haeckel E. 1879: 339 |
Solmundus tetralinus
| Mayer A. G. 1910: 452 |
| Maas O. 1905: 71 |
| Haeckel E. 1879: 351 |