
Pieza sinclairi, Evenhuis, Neal L., 2002
|
publication ID |
https://doi.org/ 10.5281/zenodo.156131 |
|
DOI |
https://doi.org/10.5281/zenodo.6277373 |
|
persistent identifier |
https://treatment.plazi.org/id/CE438843-FFAD-8814-AE24-1856FD5AB47C |
|
treatment provided by |
Plazi |
|
scientific name |
Pieza sinclairi |
| status |
|
Pieza sinclairi , sp. nov.
Mythenteles sp. McMullen, 1993: 98. McMullen & Naranjo, 1994: 26, 27.
Male. Length: 0.841.50 mm. Head ( Fig. 6 View FIGURES 5 6 ). Black to brown, overlaid with slight brownishgrayish pollen; eyes dichoptic, separated at vertex by 2 distance between lateral ocelli; front brown to yellow, depressed medially with dark brown to black medial spot; face yellow to brown; antennae ( Fig. 1 View FIGURES 1 2 ) brown to black, scape obconical, pedicel subspherical, slightly wider than long; first flagellomere ovoid, slightly longer than wide; second flagellomere short, almost half length of first flagellomere, with subapical style, apical portion of second flagellomere very thin; occiput with paired silvery grayish pollinose diadema; mentum distinctly bilobed, brownish laterally adjacent to posterior eye orbits, yellow to white along medial groove; proboscis dark brown to black, labrum pointed, length about equal to twothirds head height, proboscis able to vary in length, extendable and retractable; palpi long, thin, dark brown, extending slightly beyond length of labrum.
Thorax. Mesonotum dark brown to brown, without vittae; humeral callus, interhumeral triangle, notopleural mark extending posteriorly to prealar suture, postalar callus, small spot above postalar callus, and scutellum yellow, with scattered pale hairs on mesonotum, darker hairs on scutellum; pleura brown on lower portions of sclerites, yellow to white above; anepimeron almost all yellow; legs dusky to yellow; apical tarsal segments dark brown to black; halter stem and knob yellow to white.
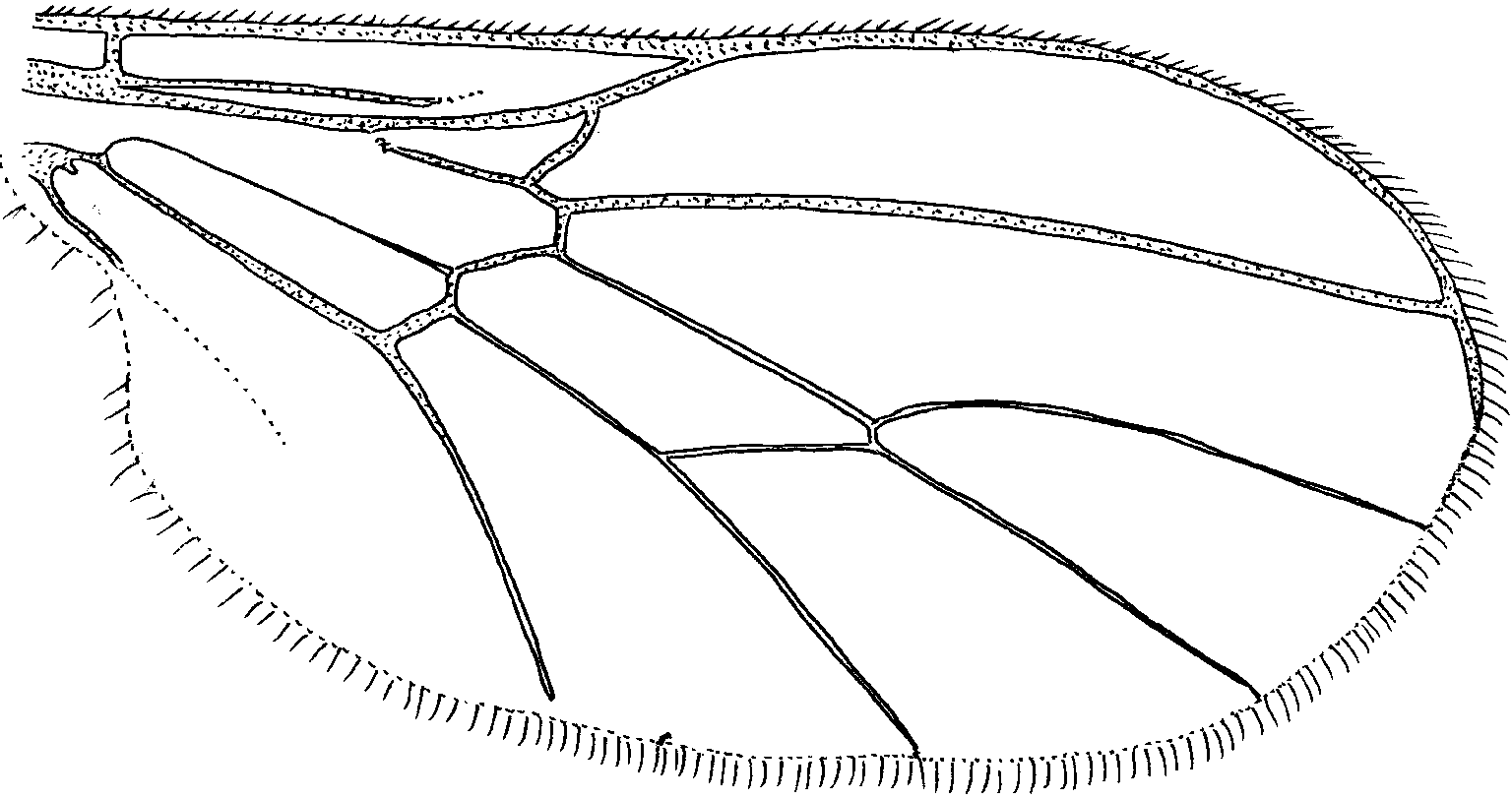
Wing ( Fig. 22 View FIGURE 22 ). Subhyaline, slightly tinged with brown color, short, stubby when compared with the congeners; veins brown; costa ends slightly before midpoint between ends of veins R4+5 and M1; vein Sc incomplete, ending at level about 1/4 distance from origin of Rs to end of R2+3; Rs evanescent at junction with R1; R2+3 arising approximately threefourths distance from origin of Rs to rm crossvein; R4+5 slightly curved to wing margin; rm crossvein at basal 1/4 of discal cell; cell dm narrowed apically; vein at base of cell m1 shorter than rm crossvein; vein M1 curved toward wing margin; vein M2 straight to wing margin; vein A1 thick only at base, evanescent for most its length to wing margin; fringe of hairs along posterior margin of wing short, fairly dense.
Abdomen. Dorsum brown, some specimens with yellow laterally on tergites; pale yellow to whitish fasciae, if present, restricted to narrow bands on posterior margins of segments, rarely found on basal segments; venter yellow, semimembranous.
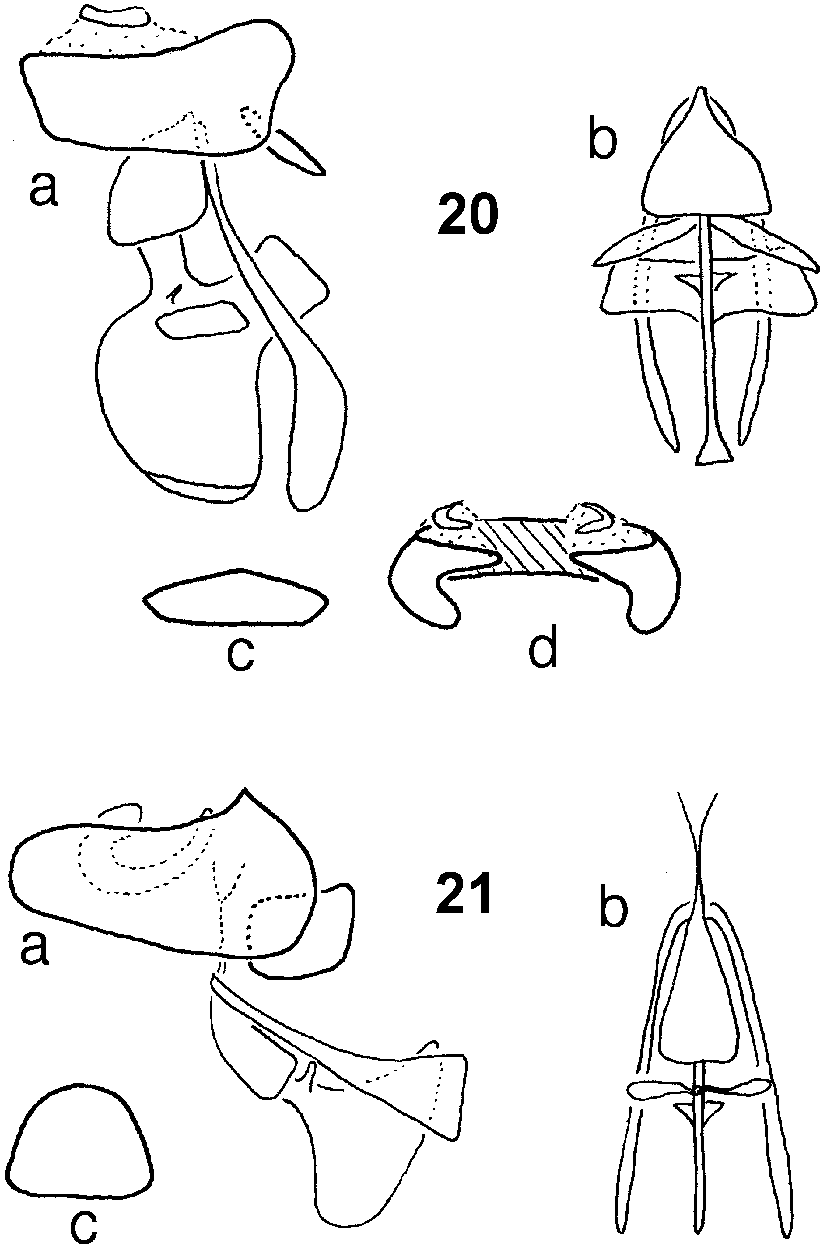
Genitalia ( Fig. 20 View FIGURES 20 21. 20 ). In lateral view with gonocoxite small, thin, subrectangular, subtriangular, rounded in dorsal view; gonostylus vestigial, fused to gonocoxite, not articulating; gonocoxal apodemes long, thin near aedeagus, flaring distally to broad, spatulate apex, darkly sclerotized; aedeagus with basal bulb shieldshaped, darkly sclerotized; aedeagal tip small, short; aedeagal apodeme large, rounded ventrally, squared dorsally, darkly sclerotized, keeled apically in ventral view, with large, subrectangular lateral vanes and another setof tiny triangular vanes; lateral rami large, foliateelongate, darkly sclerotized; epandrium long, subrectangular, pseudosurstylus well developed, pointed, incurved; cercus crescent shaped in caudal view, well exserted.
Female as in male except as follows: genitalia ( Fig. 19 View FIGURES 17 19 ) with vaginal furca Oshaped with opening at 1 end, lateral processes thick, with darkly sclerotized, thin, basal sclerites angled outward, medial processes slightly curved inward; apical process notwell developed, consisting of 2 sclerites joined medially; spermathecal reservoir somewhat peanutshaped, length about half length of sperm pump, darkly sclerotized, with subapical gland hairs in transverse band; membranous, minute, saclike area with gland hairs basal to reservoir, leading to apical spermathecal duct; apical duct extremely long, thin, membranous, length about 2.5 length of sperm pump (reservoirs situated at level of tergite II in situ); sperm pump membranous, subcylindrical; apical valve flared, with thickened and recurved outer walls; basal and common ducts not evident; vaguely sclerotized crescentshaped sclerite at base of sperm pump.
Dissection of the abdomen of a gravid female to examine the genitalia revealed 15 ovariole eggs filling the entire abdominal cavity, each about 0.07 mm long and ovoidspherical in shape, without chorional texture or pattern.
Types. Holotype male and allotype female from ECUADOR: GALÁPAGOS: Santa Cruz: Academy Bay, ECCD, 10.v14.vii.1985, 30 m, arid zone, thornscrub, MalaiseFIT trap,S. & J. Peck, deposited in CNC. Paratypes: GALÁPAGOS: Santa Cruz: 111 males and females, topotypic, collected with types ( CNC, CAR, BPBM, ECCD, USNM, BMNH); 1 male, Los Gemelos, 3 km N. Santa Rosa, 570 m, 13.vi.1985, Scalesia forest litter, S. & J. Peck ( CAR). Española: 39 males and females, Bahia Manzanilla, 510.vi.1985, littoral Cryptocarpus & Prosopis , FIT, Malaise, S. & J. Peck ( CAR). Floreana: 14 males and females, Black Beach, 2128.iii.1989, 10 m, littoral, arid, FIT, Peck & Sinclair ( CAR). Pinta: 2 females, 9.vii.1990, on flowers of Scalesia baurii, C.K. McMullen ( USNM). San Cristobal: 34 males and females, Punto Bazquero, south beach, 1421.ii.1989, littoral, yellow pan traps, B.J. Sinclair ( CAR, BPBM). Santa Fe: 1 female, 5.iv.1989, sweeping vegetation near beach, B.J. Sinclair ( CAR). Santa Isabela: 16 males and females, 4 km NW Villamil, 215.iii.1989, arid forest, 20 m, FIT, Peck & Sinclair ( CAR). CURAÇAO: 15 males and females, Coral Specht, 3 km E. Willemstad, 915.ii.1997, dungbaited pitfall trap in desert scrub near coast, W.E. Steiner & J.M. Swearingen ( USNM). DOMINICAN REPUBLIC: 1 male, 1 female, Pedernales, 14.5 km N. Cabo Rojo, 165 m, 18° 03' N 71° 39' W, 13–19.vii.1990, L. Masner, J. Rawlins, C. Young, arid thornscrub, intercept trap ( CMNH). The specimens from Floreana and Santa Isabela are preserved in alcohol, the remainder are preserved on pins.
Etymology. The species is named for Brad Sinclair, who has significantly improved our knowledge of the Galápagos fly fauna.
Remarks. Greasy specimens have the interhumeral, prescutellar, and notopleural marks vague, and the color of the thorax and abdomen may appear dark brown to black. Fresh specimens are all dusky brownish with pale pubescence. The scutellum can vary from all yellow, yellow with a median black spot, to brown with yellow only on sides. The female spermathecal reservoir is very similar in some respects to that of Paraconsors (Paraconsors) scutellaris (Melander) from the southwestern United States, though the two species are not related.
Specimens have been collected from 13 February to 14 July. Due to its being found in relatively tropical latitudes where seasonality in bombyliids is uncommon, it could be expected that this species would be found all year long. McMullen (1993) recorded this species (as “ Mythenteles sp.”) from flowers of the composite Scalesia baurii in the Galápagos.
This species is the most common of all the species of Pieza thus far collected, which no doubt only reflects the poor concentrated collecting for other species. Specimens of these species were collected using a mixture of collecting trap methods, but the most successful appear to be the flight intercept traps (FIT) and pan traps. The majority of the specimens collected in this fashion consisted of females. This phenomenon was also found in insular specimens of Empidideicus freyi Greathead. Greathead (1986) surmised that this disparate sex ratio could be due to swarming behavior.
Distribution. Known only from a relatively small series of specimens from Curaçao (15) and the Dominican Republic (2) and the large series of specimens from the Galápagos. It is doubtful that the Galápagos population is indigenous to the islands. Previous records of collecting there ( Coquillett 1901; Curran 1932, 1934; Johnson 1924; Linsley and Usinger 1966) do not mention microbombyliids and, given that sinclairi has been collected by the 100s in Malaise and other insect traps in the 6 Galápagos islands listed above (and many 100s more unmounted specimens exist in CAR and CNC), it is highly probable that this species represents a recent accidental introduction. However, until the immature stages of sinclairi are known, it is impossible to say for sure how it could have been introduced. Adult are weak fliers, fly close to the ground and, therefore, are not good dispersers. The nonGalápagos localities of Curaçao and the Dominican Republic, both insular habitats, could also be evidence of introduced populations of this species. It is possible that concentrated collecting in Colombia, Venezuela, and the Ecuadorian mainland could turn up more specimens of this species.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pieza sinclairi
| Evenhuis, Neal L. 2002 |
Mythenteles
| McMullen 1994: 26 |
| McMullen 1993: 98 |