
Vegaranina rivasi, Arano-Ruiz & Viñola-López & Rojas-Consuegra & Borges-Sellen, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4527.4.10 |
|
publication LSID |
lsid:zoobank.org:pub:B366D5D3-4F14-41B9-94F5-6FB28A9784AB |
|
DOI |
https://doi.org/10.5281/zenodo.5996670 |
|
persistent identifier |
https://treatment.plazi.org/id/CD71E14A-FF9A-FFF8-FF07-FE803894FBDF |
|
treatment provided by |
Plazi |
|
scientific name |
Vegaranina rivasi |
| status |
sp. nov. |
Vegaranina rivasi View in CoL sp. nov.
( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Lophoranina precocious, Varela & Rojas-Consuegra 2009: 118 View in CoL , 119, 122; Varela & Rojas-Consuegra 2011: 3, 6; Van Bakel et al. 2012: 95; Luque et al. 2017: 56. [Not L. precocious Feldmann, Vega, Tucker, Garcia-Barrera & Avendano, 1996 View in CoL ]
Material examined. The holotype (MMC- 5-68-1) is an almost complete carapace, preserving all the terraces, anterolateral spines, extraorbital tooth and part of the tridentate rostrum. The specimen has a diagonal fracture from the right margin of the 4 th terrace to the midline of the branchiocardiac groove. The paratypes (MMC- 5-68-2, 5-68- 3) are right and left half of the carapaces of two specimens partially preserving some of the terraces. The specimen MMC- 5-68-1 was collected in 1999 by Luis Orlando, and the paratypes were collected by Jesús Servilio Quintero in 2014. All specimens are deposited at the Museo Municipal de Cruces (MMC).
Measurements (in mm): Holotype MMC- 5-68-1 length: 90, width: 75; paratype MMC- 5-68-2 length (estimated) 50, width (estimated) 40.
Type locality. The holotype (MMC- 5-68-1) was collected about 1.5 km south of the town Potrerillo, while the paratypes (MMC- 5-68-2, 5-68-3) at 1 km east of the same town, in the Municipality of Cruces, Cienfuegos Province, Cuba.
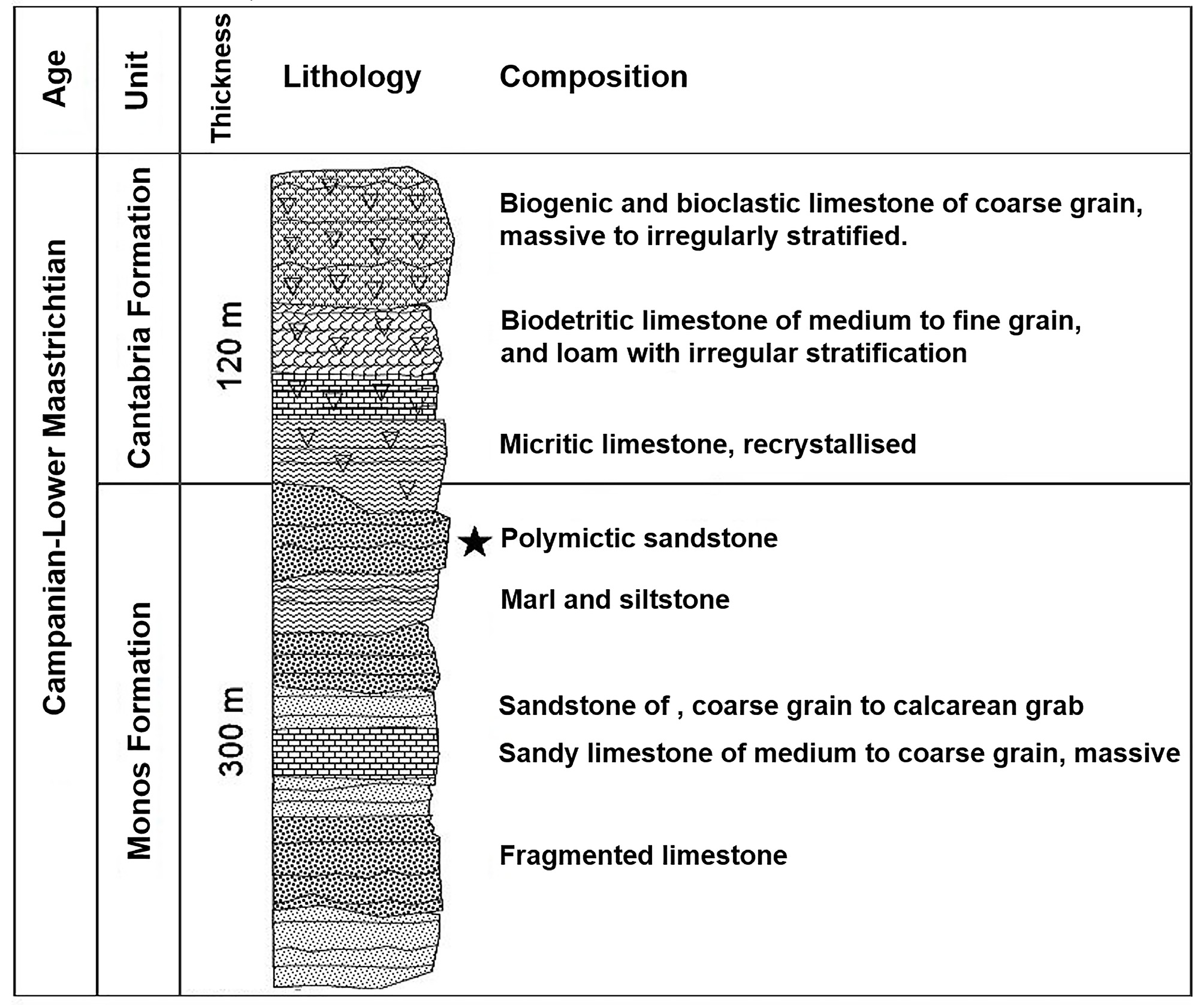
Geologic formation and age. Recent investigations of the area of provenance of the specimens indicates that those rocks belong to the Monos Formation (Truitt, P., in Brönnimann & Pardo 1954) and not the Cotorro Formation as was initially suggested by Varela & Rojas-Consuegra (2009). The Monos Formation represents a transgressive cycle characterized by a sequence of polymictic gravels that transgress into sandstones, formed by the erosion of the Antillean Cretaceous Volcanic Arc ( Fig. 1 View FIGURE 1 ). Limonites, and loam can also be found intercalated in some sections of the outcrop. These rocks were deposited during the Campanian - Lower Maastrichtian in a shallow marine environment with strong input of terrestrial sediments that was decreasing over time as the transgression progressed (IGP, 2013). The three specimens were found in sandstone nodules weathered out from the structural rock. Several ammonites, gastropods, bivalves, echinoderms and wood fragments have also been collected from the same sandstone section. In the region, the Monos Formation is transitionally covered by Maastrichtian carbonates of the Cantabria Formation (Truitt, P., in Brönnimann & Pardo 1954; Kantshev et al. 1976).
Etymology. The species is named in memory of Danilo Rivas Enterrio (1962–2014) who was an important collector of fossils from Cienfuegos, Cuba.
Diagnosis. Species of Vegaranina , with the following unique combination of characters: large anterolateral spines (larger than post-orbital spine), lateral and posterolateral margin with small spines. First and second transverse terraces plus mesogastric, metagastric, and cardiac region fused at carapace midline ( Fig. 2 View FIGURE 2 , 3 View FIGURE 3 ); interrupting cervical, cardiac and branchiocardiac grooves. Anterior and posterior terraces of branchial region partially fused.
Description. Large, wide raninid, with ovoid carapace, convex transversely, reaching its maximum width at level of middle epibranchial terrace. Carapace converging strongly towards the posterior region, from fourth transverse terrace posterior to the cardiac region. Lateral and post-lateral margin of the carapace serrated with small spines ( Fig. 4 View FIGURE 4 ), similar to serration on terraces. Carapace divided into three main anteroposterior sections: rostrum and post-frontal region; lobulated terraces between the postfrontal region and branchiocardiac groove; seven transverse terraces from side to side posterior to branchiocardiac grooves. Tridentate rostrum not completely preserved, axial spines serve as inner orbital spines, right lateral spine from the rostrum preserved. Orbits wide and concave, with two large and open fissures; inner one placed between inner orbital spine and blunt intra-orbital spine. Outer fissure located between blunt intra-orbital spine and outer orbital spine, which is directed anteriorly. Anterolateral margin bears three prominent and flattened spines, longer than wide, larger than post-orbital spine, directed anterolaterally and anteriorly depending on the spine. Post-frontal region broad and non-terraced. Lobulated terraces separated from the post-frontal region by cervical grove, and from the seven main transverse terraces by branchiocardiac. Lobulated terraces and the ones located posterior the branchiocardiac groove, are serrated on the anterior ridge and gently inclined backward. Epibranchial region with three serrated terraces. Epibranchial region separated from branchial and metagastric region by a diagonal groove. Branchial region composed of two terraces nearly completely fused, anterior terrace near five times smaller than posterior terrace. Branchial region separated from cardiac and metagastric region by diagonal groove. Mesogastric, metagastric and cardiac region plus first and second transverse terraces fused on the midline by a stripe 5 mm wide. Fusion of previous mentioned structures interrupt the cervical, cardiac, and branchiocardiac grooves. Cardiac lobes appear to be rhomboidal and not serrated. Region posterior to branchiocardiac groove and composed by metabranchial, mesobranchial, and intestinal region are undifferentiated; cut by seven strong serrated terraces transverse from side to side. Terraces are concave and resembles a wide U. Other terraces show lower level of fusion or connection, varying among individuals.
Remarks. The genus Vegaranina was erected by Van Bakel et al. (2012) to contain Lophoranina precocious and two undescribed species. Vegaranina rivasi sp. nov, differs from V. precocia by the fusion of the mesogastric, metagastric and cardiac region as well as the first and second transverse terraces on the midline by a stripe of 5 mm. In V. precocia , each of these structures is separated by deep grooves. The lateral margins of the carapace of V. rivasi sp. nov are serrated, but these are smooth in V. precocial . The three anterolateral spines are larger in V. rivasi sp. nov., although Feldmann, and Schweitzer (2007) observed that the length of the spines could vary because of sexual dimorphism. Among the lobulated terraces, the cardiac terraces of V. rivasi sp. nov. are subrhomboidal, while in V. precocia they tend to be rectangular. The differences observed between the specimens from Cuba and Mexico cannot be explained by ontogenetic changes because the characters that distinguish the species are stable in individuals of distinct size in each taxon ( Feldmann et al. 1996, Vega et al. 2018); also, the size of the paratypes of the Cuban species (MMC- 5-68-2, 5-68-3) overlap with the size of V. precocia . Although the sex of the specimens from both species has not been determined, it seems unlikely that the seven fossil specimens of V. precocia reported by Vega et al. (2018) represent one sex and the three Cuban specimens the other. We consider that with the exception of the differences in spine size, sexual dimorphism fails to be the cause of the difference between other characters. Lastly, Klompmaker et al. (2015) demonstrated that the differential preservation of the cuticle in fossil decapod crustaceans can have important effects for species identification. Nevertheless, in all specimens of V. rivasi sp. nov., and in the holotype ( Feldmann et al. 1996, fig. 3) and neotype of V. precocia , ( Vega et al 2018, fig. 6) the cuticle is preserved.
Schweitzer et al. (2008) reported Lophoranina cf. precocious from the Late Cretaceous of Puerto Rico. However, the specimen lacks most of the region anterior to the branchiocardiac groove, and the characters that differentiate V. precocia and V. rivasi sp. nov. cannot be evaluated. We recommend to referring the Puerto Rican specimen to Vegaranina sp. until more and better preserved specimens are found.
Feldmann et al. (1996) proposed that the presence of fewer and stronger terraces in V. precocia ( L. precocious ) is a primitive character that evolved later in numerous, and thinner terraces in the Eocene-Miocene Lophoranina . Van Bakel et al. (2012) stated that Vegaranina could be the link between the Cenomanian Lophoraninella cretacea (Dames, 1886) and Eocene-Miocene Lophoranina . Nonetheless, L. cretacea is characterized by two anterolateral spines and several thin terraces ( Van Bakel et al. 2012), like Lophoranina ( Karasawa et al. 2014) , and it seems unlikely that in the transitional form ( Vegaranina ) these key characters would be different. Posteriorly, Karasawa et al. (2014) in a phylogenetic study of Raninoida found that Lophoranina and Vegaranina are sister genera, linked by the character of carapaces with terrace ornamentation. Nevertheless, Lophoraninella cretacea was not included in the phylogeny by Karasawa et al. (2014), because the type specimens are poorly preserved. It seems that until better specimens of Lophoraninella are found, its phylogenetic position in respect to Lophoranina and Vegaranina remains unclear.
The distribution pattern of Vegaranina spp. in America and Europe during the late Cretaceous support the hypothesis that the genus originated in the Tethys region ( Feldmann et al. 1996, Schweitzer et al. 2007). It also indicates that the genus was widely dispersed, with at least 3 species living throughout the Tethys ( Van Bakel et al. 2012, Vega et al. 2018).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Vegaranina rivasi
| Arano-Ruiz, Alberto, Viñola-López, Lazaro W., Rojas-Consuegra, Reinaldo & Borges-Sellen, Carlos Rafael 2018 |
Lophoranina precocious
| Luque, J. & Schweitzer, C. E. & Santana, W. & Portell, R. W. & Vega, F. J. & Klompmaker, A. A. 2017: 56 |
| Van Bakel, B. W. M. & Guinot, D. & Artal, P. & Fraaije, R. H. B. & Jagt, J. W. M. 2012: 95 |
| Varela, C. & Rojas-Consuegra, R. 2011: 3 |
| Varela, C. & Rojas-Consuegra, R. 2009: 118 |