
Sparasion Latreille
|
publication ID |
https://doi.org/ 10.1206/578.1 |
|
persistent identifier |
https://treatment.plazi.org/id/CB538789-546F-3141-D20C-BEC2FC35FDC2 |
|
treatment provided by |
Carolina |
|
scientific name |
Sparasion Latreille |
| status |
|
Sparasion Latreille View in CoL
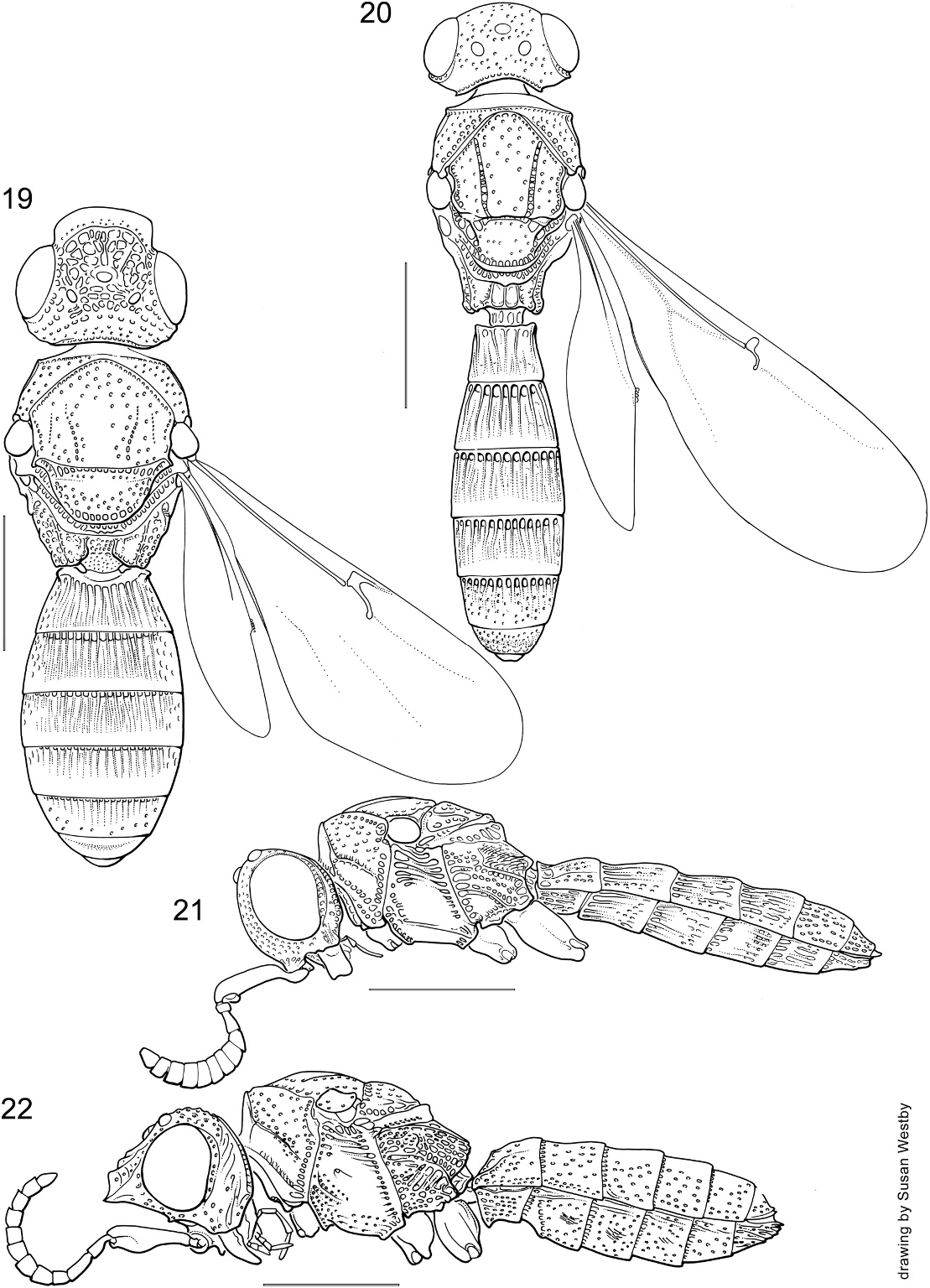
Figures 19, 22 View Figs , 24 View Figs , 35 View Figs –40
Sparasion Latreille, 1802: 316 View in CoL . Type: Sparasion cephalotes Latreille View in CoL , by monotypy.
Oxyurus Lamarck, 1817: 128 View in CoL . Type: Sparasion frontalis Latreille , designated by Muesebeck and Walkley (1951). Preoccupied by Oxyurus Rafinesque, 1810 (Pisces) View in CoL .
Bebelus Gistel, 1848 : x. Type: Sparasion frontalis Latreille , by substitution of Bebelus for Oxyurus View in CoL . Replacement name for Oxyurus Lamarck. View in CoL
Prosparasion Kieffer, 1913: 190 . Type: Prosparasion coeruleum Kieffer , by monotypy and original designation. Synonymized by Masner (1976).
DESCRIPTION: Small to large, length 2.1– 7.3 mm; body robust (figs. 19, 22, 35), sometimes slightly depressed, with relatively long, strong legs, antenna relatively short, width of antennomeres variable; head large, usually with one to three transverse ledges in upper portion of frons; head, body usually black, sometimes with metallic blue or green tinge, sometimes brown, yellow, or bright orange; macropterous.
Head large, weakly transverse to quadrate in dorsal view (figs. 19, 38); upper frons usually produced anteriorly into one to three transverse shelves (figs. 22, 35, 36), anterior margin simple, usually arcuate, sometimes sinuous; hyperoccipital carina absent; occipital carina well developed, continuous medially, crenulate; lateral ocellus distinctly separat- ed from inner orbit of compound eye, OOL 0.5–1.5 times diameter of lateral ocellus (fig. 38); compound eye glabrous or with few short setae (figs. 37, 40); frons shape variable, weakly concave to broadly convex, without distinct scrobe, sculpture highly variable, usually with transverse carina above interantennal prominence (fig. 37: tc); area between compound eye, antennal insertion and base of mandible deeply concave corresponding to shift in position of anterior mandibular articulation; interantennal process strongly produced anteriorly, torulus opening laterally from process; submedian carina absent; orbital carina absent; lower frons without fanlike striae; interocular space broad, subequal to eye height; inner orbits parallel; clypeus strongly raised beneath interantennal process, elongate, apically truncate or rounded, not divided into anteclypeus and postclypeus; malar sulcus absent; gena moderately expand- ed, convex; unsclerotized labrum, apical fringe of long setae often exposed; mandible large, lower margin distinctly expanded so that mandible is widened toward apex, anterior articulation deeply invaginated, posterior articulation shifted anteriorly to side of head, mandibular range of movement primarily anteroventral; mandible usually with two, rarely three apical teeth, longitudinal axis of mandible slightly twisted so that teeth approximately transverse; maxillary palpus 5- segmented, segment 3 expanded medially; labial palpus 3-segmented; antenna 12-merous in both sexes; radicle inserted subapically into A1, nearly perpendicular to longitudinal axis of A1 (fig. 37: r); A1 distinctly widened in apical half; A3 longer than A2; female antenna without distinct clava; gustatory sensilla on female antenna arranged in longitudinal pairs on apical antennomeres; claval formula A4–A12 1-2-2-2-2-2-2-2-1; male antenna with tyloids on A4–A11, sometimes A12.
Mesosoma robust, in dorsal view longer than wide (figs. 19, 38), in lateral view deep, somewhat flattened dorsally (figs. 22, 36, 40); pronotum in dorsal view broad laterally, anterolateral corners angulate; transverse pronotal carina present (fig. 38: tpc), sometimes effaced medially; vertical epomial carina variably developed, usually present dorsally; horizontal epomial carina present, strongly developed; anterior face of pronotum short, but clearly visible in dorsal view; lateral face of pronotum broadly concave below horizontal epomial carina, facing anterolaterally, with broad scrobe for reception of foreleg; netrion apparently absent, posterior margin of lateral pronotum bordered by deep longitudinal costae; anterior margin of mesoscutum meeting pronotum dorsally; mesoscutum triangular to pentagonal in outline, posterolateral corner produced into acute flange flanking tegula (fig. 38: f); parapsidal lines present; notauli variably developed, sometimes absent, often obscured amid coarse mesoscutal sculpture; skaphion absent; transscutal articulation well developed, crenulate to costate; scutellum wider than long, unarmed laterally, convex to weakly flattened; axilla small; metanotum narrow, dorsellum clearly differentiated, armature variable, sometimes produced medially into weak spine; dorsal surface of propodeum with dense, fine pilosity; keels, plicae of propodeum variably developed; posterior face of propodeum punctate, setose; mesopleuron large, prominent; mesopleural depression well developed (fig. 36: dep); anterior margin of depression sharply demarcated by change in sculpture, but without distinct mesopleural carina; sternaulus absent; mesopleural pit present, shallow; anterior margin of ventral portion of mesepisternum and acetabular carina extended forward moderately between forecoxae; mesepisternum and mesepimeron separated by line of well developed foveae; episternal foveae absent; dorsal corner of mesepimeron produced into short, acute, posterior tooth; anteroventral portion of metapleuron separated from lateral face by irregular carina and sculpture, setose; metapleural pit present near anterior margin of metapleuron; posterior margin of metapleuron lamellate; metapleuron separated from propodeum dorsally by deep groove; propodeum with longitudinal carinae variably developed, setose throughout, posterolateral corners projecting posteriorly as blunt prominences; legs robust (fig. 35), femora weakly to distinctly incrassate; posterior surface of hind coxa with arcuate striae, setose; trochantellus absent from all legs; outer surface of fore-, midtibiae with strong semierect spines and elongate setae; tibial spur formula 1-2-2, inner spur longer; tarsal formula 5-5-5; tarsomeres tapering in width apically; pretarsal claws simple; apex of forewing extending to T6 or beyond, variably infuscate, marginal cilia short, but distinct; R fairly straight (figs. 19, 39), extending through basal 0.5 of length of forewing, interrupted by distinct bulla basad of origin of r-rs, with strong bristles arising throughout its length; R 1 usually reaching costal margin, extending apically variable distance as postmarginal vein, vein inflated posteriorly and costal cell posterior to R 1 deeply pigmented, forming pseudostigma; r-rs (stigmal vein) reflexed apically, arising just beyond bulla in R, away from costal margin; no other tracheate veins in forewing; hind wing with R tracheate only at extreme base, not extending to hamuli and costal margin; no strong dark bristles on R; costal margin of hind wing above R with rather dense, strong setae; four hamuli present.
Metasoma more or less cylindrical (figs. 19, 22, 35), terga slightly flattened, sterna deep, convex; T1–T5 subequal in length, T1 slightly the longest; female with six terga, six sterna visible externally, male with eight terga, seven sterna visible externally; submarginal ridge well developed, defined by narrow laterotergites to form deep submarginal rim; no spiracles visible; base of segment 1 longitudinally costate; suture between segments 1 and 2 basally crenulate; base of segments 2–6 costate, sutures between segments beyond 6 simple; female T6 without median raised field of microsetae or secretion; S1 not laterally compressed, clasped between apices of hind coxae, not extending anteriorly between bases hind coxae; anterior margin of S2 straight; no felt fields present on sterna; S7 with lateroventral apodemes; T7 not extruded with ovipositor, with short internal paired apodemes; S7 with paired lateral apodemes.
DIAGNOSIS: The combination of a transverse ledge on the frons, 1-2-2 tibial spur formula, and presence of a bulla in R is found elsewhere only in Listron , new genus. Sparasion may be distinguished from it by the presence of a differentiated radicle, the expansion of the ventral margin of the mandible, the expanded third maxillary palpomere, and the shift in position of mandibular articulations with its corresponding effect on the shape of the lower frons and clypeus. The few species of Sparasion lacking a frontal ledge, e.g., S. coeruleum Kieffer (fig. 40), may be distinguished from Mexon new genus by the broad LOL, and from Sceliomorpha by the differentiated radicle.
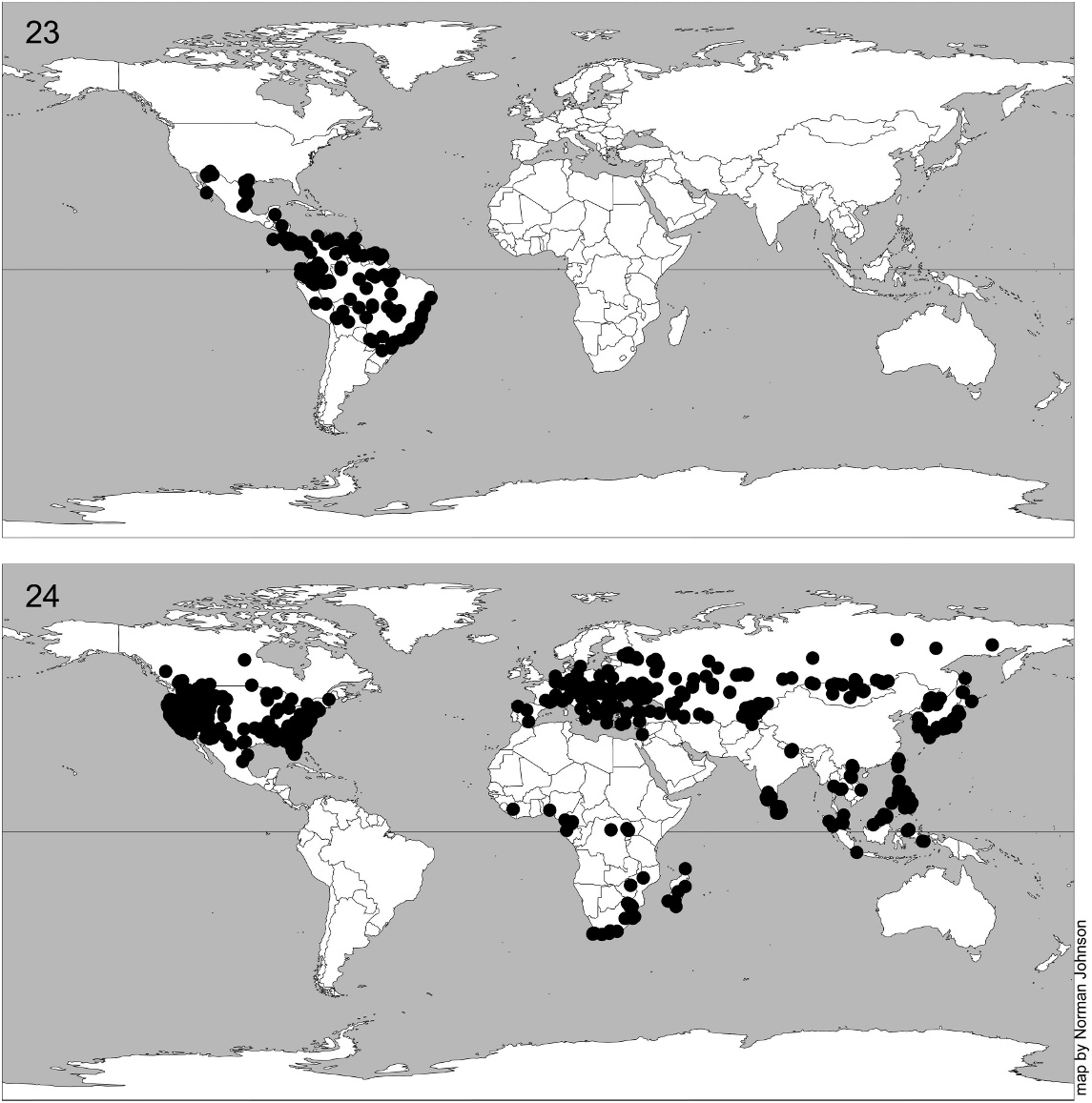
GEOGRAPHIC DISTRIBUTION: Widespread in Eurasia, Africa, and temperate North America (fig. 24). Unknown from Australasia; a single specimen is known from Argentina in CNCI, possibly either an introduction or a mislabelled specimen. Two species also recorded from Baltic amber ( Brues, 1940).
HOSTS: Recorded in the United States from Anabrus simplex Haldeman and Atlanticus gibbosus (Scudder) ( Muesebeck, 1979; Grissell, 1997) ( Orthoptera : Tettigoniidae , Tettigoniinae ).
COMMENTS: The holdings of the Muséum National d’Histoire Naturelle in Paris includes several specimens, generally in poor condition, bearing labels from the Bosc collection suggesting that they may have been in the material studied by Latreille (Notton, personal commun.). None, however, match the description and provenance of the two relevant type species here, Sparasion cephalotes and S. frontale . Although we thus cannot document completely the identity of Sparasion and its synonym Oxyurus (and Bebelus ) through the primary types of the type species of the genera, this is of little practical import. Sparasion has been consistently recognized for over 150 years and, in the European fauna, it is unmistakable.
Kieffer (1913) described the genus Prosparasion , distinguishing it from Sparasion solely on the basis of the lack of a transverse carina on the frons (fig. 40). Masner (1976), without elaboration, synonymized the two names. Kelner-Pillault (1958) earlier claimed that the holotype is deposited in the MNHN, but this specimen comes from Butuan on the island of Mindanao, not Luzon. Therefore it cannot be the holotype. Much of the Baker collection is now found in the National Museum of Natural History, Washington, DC. Among these is one specimen, bearing a type label in Kieffer’s hand, from ‘‘Los Banos, P. I. ’’ While this is not literally the type locality reported by Kieffer in the original description, ‘‘Mt. Makiling’’, the city of Los Baños is at the foot of the mountain. We have found no specimens in any of the collections that have Mt. Makiling on the label. Accordingly, we conclude that the NMNH specimen is the holotype of Prosparasion coeruleum .
Prosparasion coeruleum is very distinctive (fig. 40): The frons is evenly convex and lacks any indication of a transverse ledge. Almost all other species of living Sparasion View in CoL possess such a ledge, sometimes even two or three. The vein r-rs in the forewing is nearly parallel with the costal margin. The metallic blue color is not unique to this species nor particularly uncommon in the genus, but is a notable departure from the usual black to dark brown color found in Sparasion View in CoL . On the other hand, P. coeruleum shares with all extant Sparasion species the medial posterior flangelike expansion of the mandible and the presence of an irregular transverse carina directly above the antennal insertions. These last two characters are unique to extant Sparasion View in CoL . In the absence of a worldwide revision of Sparasion View in CoL , we believe that it is both prudent and conservative to support the synonymy originally proposed by Masner (1976). This synonymy of generic names renders Prosparasion coeruleum Kieffer, 1913 a junior homonym of Sparasion coeruleus Kieffer, 1905 . Accordingly, we propose the replacement name, Sparasion lividus Johnson, Masner and Musetti new name for the type species of the genus Prosparasion .
Uncertainty concerning the gender of the name Sparasion View in CoL has led to confusion as to the proper endings to use with specific epithets. In his original description of the genus, Latreille (1802) included only a single species, S. cephalotes Latreille View in CoL , an epithet that does not distinguish among the three possibilities of masculine, feminine, and neuter. Three years later ( Latreille, 1805), he added a second species, S. frontalis Latreille. This suggests that the gender of Sparasion View in CoL is not neuter. However, in the next publication in which he treated the genus, Latreille (1809) emended the ending of this second species to S. frontale . This seems to be strong evidence that, at least at that time, he considered the name to be neuter.
The name Sparasion was generally, but not unanimously, used as a neuter noun through the rest of the 19th and into the 20th century. The change of usage to the masculine appears to have begun with Kieffer (1916), but probably received its greatest impetus by the nearly systematic change of specific epithets in his compendium of the world Scelionidae ( Kieffer, 1926) . This usage has now been followed in the description of the many species from the Palearctic ( Kozlov and Kononova, 1988, 1990; Kononova, 1995; Kononova and Petrov, 2001a, 2001b). Given the general confusion on this issue, any decision will result in the need to change a good number of names from one gender to the other. Therefore, because Latreille (1805) first used a nonneuter epithet ( frontalis ) and the name has never been considered to be feminine, we believe it is best to follow Kozlov and Kononova (1988, 1990) and treat Sparasion as a masculine noun.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Sparasion Latreille
| Johnson, Norman F., Masner, Lubomír & Musetti, Luciana 2008 |
Prosparasion
| Kieffer, J. J. 1913: 190 |
Oxyurus
| Lamarck, J. B. 1817: 128 |
Sparasion
| Latreille, P. A. 1802: 316 |