
CREODONTA
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00194.x |
|
persistent identifier |
https://treatment.plazi.org/id/CB5187EB-FFB6-FF89-4D04-FD53BF52A004 |
|
treatment provided by |
Diego |
|
scientific name |
CREODONTA |
| status |
|
MACHAEROIDINAE, CREODONTA
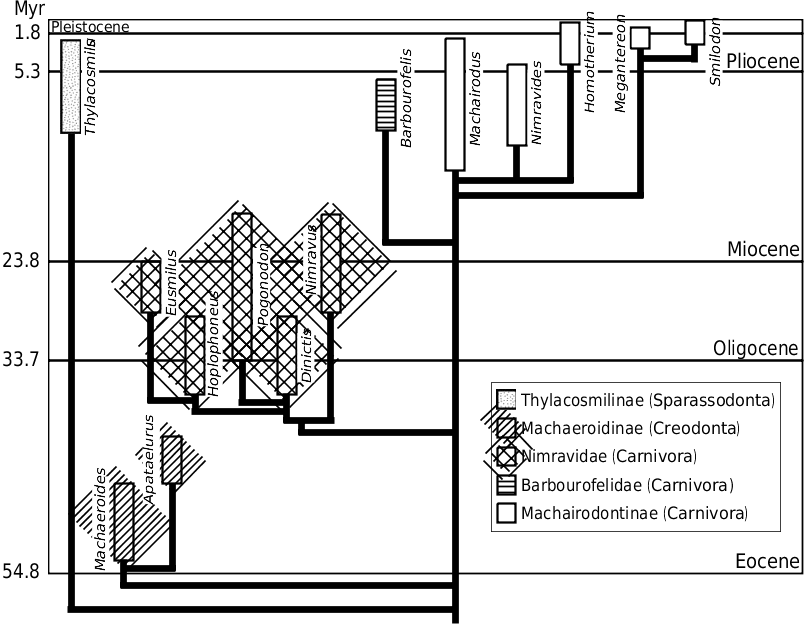
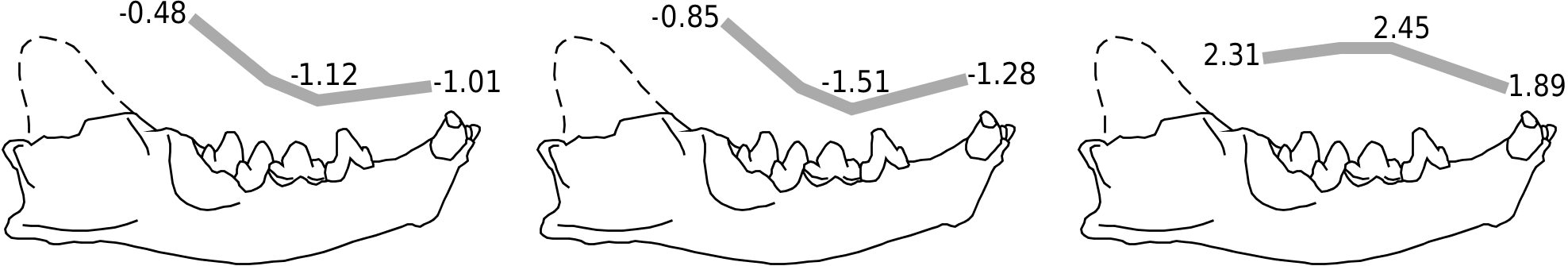
Machaeroidines are known from lower to middle Eocene deposits of North America ( Fig. 1 View Figure 1 ). They differ dentally from carnivorans in having the second lower molar (rather than the first) modified into a carnassial (see Gunnell, 1998). Only two machaeroidine specimens have mandibles sufficiently complete for detailed analysis: the small Machaeroides eothen (USNM 17059; see Gazin, 1946) and the larger Apataelurus kayi (CM 11920; see Scott, 1938). Even though the mandibular flange of Apataelurus kayi is only partially preserved, the mandibular symphysis is complete and allows for an accurate determination of mandibular depth at the canine. Additionally, partial mandibles for Machaeroides eothen (USNM 361372) and the slightly more derived and younger Machaeroides simpsoni (CM 36397; see Dawson et al., 1986) were studied. Lacking the postcarnassial segment of the ramus, mandibular force profiles could not be evaluated for these specimens because accurate determination of the moment arm length for each interdental gap is impossible. However, the bending strengths at each interdental gap, Zx and Zy, and relative mandibular force (Zx / Zy) could be calculated and compared to those of the complete specimen of M. eothen . By doing so, intrageneric variability can be evaluated, providing insight into possible phylogenetic trends.
Mandibular force profiles
Although Machaeroides eothen and Apataelurus kayi have slightly different mandibular force profiles, both exhibit a negative slope in dorsoventral (Zx / L) and labiolingual (Zy / L) forces between M 1 M 2 and P 3 P 4 and a very strong symphyseal region, much stronger than the corpus at the carnassial ( Fig. 5A View Figure 5 ). The peak in dorsoventral force at P 4 M 1 in M. eothen is an artefact produced by the anomalous expansion of the dorsal margin of the mandibular corpus into the interdental gap between the right P 4 and M 1 in USNM 17059, artificially increasing mandibular depth at that interdental gap.
Comparison of the dorsoventral and labiolingual bending strengths (Zx and Zy) for the two specimens of M. eothen and M. simpsoni confirms the anomalous mandibular properties at P 4 M 1 in USNM 17059 ( Fig. 5B View Figure 5 ): while the dorsoventral bending strength of the mandibular corpus varies little along the tooth row in the incomplete specimens of M. eothen and M. simpsoni , the Zx values at P 4 M 1 are much higher in USNM 17059. This anomaly is not as apparent labiolingually, where the bending strength values increase slightly anteriorly in both specimens of M. eothen ; in contrast, the bending strength values decrease slightly anteriorly in M. simpsoni ( Fig. 5B View Figure 5 ).
The relative mandibular force (Zx / Zy) profiles of all machaeroidine specimens studied ( Fig. 5A–B View Figure 5 ) can be compared. Omitting the exaggerated P 4 M 1 value in USNM 17059, the Zx / Zy profiles of M. eothen and A. kayi show a slow, linear decrease anteriorly between the carnassial and the canine. In contrast, a gradual increase in Zx / Zy values is observed in M. simpsoni . All machaeroidines attain minimum Zx / Zy values at the canine: Zx / Zy canine values for M. eothen vary between 1.33 and 1.60 (mean of 1.46 when mandibular dimensions are averaged prior to determination of section moduli), Zx / Zy canine = 2.15 for M. simpsoni , and Zx / Zy canine = 1.48 for Apataelurus kayi .
Interpretation
The dorsoventral and labiolingual force profiles of machaeroidines give insight into their bite force ( Table 1). At the carnassial (M 1 M 2), the bite force of Apataelurus kayi was approximately 3.50 times greater than that of the smaller Machaeroides eothen . By comparing these results with those obtained for extant taxa, A. kayi was likely able to generate bite forces comparable to those of Panthera pardus , a felid of similar mandibular length. Although felids of mandibular length similar to M. eothen were not studied by Therrien (2005), the bite force of the small machaeroidine appears to have been half that of Eusmilus cerebralis , a nimravid of similar mandibular length, one third that of Puma concolor , a felid with a mandible 50% longer, and close to that of Canis latrans , a canid with a mandible 63% longer.
The dorsoventral and labiolingual force profiles (Zx / L and Zy / L, respectively) also indicate that the mandible of machaeroidines is stronger at the canine than at the cheek teeth, suggesting that they delivered powerful canine bites to kill their prey, like felids, rather than shallow bites like canids and hyaenids
A Dorsoventral force Labiolingual force Relative force (Log Zx/L) (Log Zy/L) (Zx/Zy)
Machaeroides eothen (L = 8.95 cm) -0.47 2.05 1.48
Apataelurus kayi (L = 14.59 cm)
( Fig. 4). The close correspondence of the section modulus values of the incomplete M. eothen and M. simpsoni specimens to those of the complete M. eothen specimen reveals that the killing behaviour of these individuals must have been similar.
The relative mandibular force (Zx / Zy) profiles reveal that the mandible of machaeroidines, with the exception of M. simpsoni , becomes rounder from M 1 M 2 toward the canine ( Fig. 5 View Figure 5 ). These decreasing Zx / Zy values indicate that torsional stresses are more important toward the canine, most likely in response to prey capture. Machaeroides simpsoni is unique among machaeroidines in having a mandibular corpus that becomes deeper anteriorly between M 1 M 2 and P 3 P 4 ( Fig. 5 View Figure 5 ). Although none of the extant carnivorans studied by Therrien (2005) displayed this feature, the fact that some nimravids and machairodonts exhibit a similar trend in mandibular shape (see below) indicates that this feature evolved independently in different sabretooth lineages. The Zx / Zy profiles reveal that the mandible of the two Machaeroides species is better dorsoventrally buttressed than that of A. kayi . In other words, labiolingual stresses are relatively more important in the larger machaeroidine species than in the smaller species. In fact, whereas the Zx / Zy profile near the cheek teeth of A. kayi is within the range of extant felids, those of Machaeroides are slightly higher ( Figs 4, 5 View Figure 5 ). This difference could potentially reflect the fact that small machaeroidines captured small prey, which would have induced significantly less torsional stresses than larger prey, thus allowing the mandible to be better dorsoventrally buttressed than that of Apataelurus . However, beam models for extant carnivorans of small body size, similar to Machaeroides , and feeding on small prey need to be constructed to test these hypotheses.
The Zx / Zy canine values of M. eothen and A. kayi are similar to those of extant conical-toothed felids with special killing techniques ( Fig. 4). From these mandibular properties, it can be surmised that machaeroidines delivered canine bites on prey that offered relatively less resistance than those of extant, large, conical-toothed felids. Thus, prey must have been at least partially restrained before the sabre-like canines were used to kill. Although postcranial remains for A. kayi are not known, the humerus of M. eothen (USNM 17059) is robust and possesses a large deltopectoral crest reminiscent of that observed in dirk-toothed sabretooth ecomorphs ( Gazin, 1946; pers. observ.). Although Anyonge (1996) did not include machaeroidines in his study, he demonstrated that dirk-toothed ecomorphs had extremely powerful forelimbs. This interpretation is supported by the brachial index (maximum length of radius/maximum length of humerus) of M. eothen (BI = 0.76, based on measurements in Gazin, 1946), which is lower than that of Smilodon but slightly higher than those of Barbourofelis lovoerum and B. fricki (see below). Given the robust postcranium of machaeroidines, sabretoothed creodonts could have been able to at least partially immobilize prey with their forelimbs prior to delivering a canine bite. Finally, the fact that M. eothen and A. kayi possess lower Zx / Zy canine values than later sabretooths (possibly with the exception of Nimravus brachyops , see below) suggests that machaeroidines may have been less efficient at restraining their prey than later sabretooths (see Discussion).
Machaeroides simpsoni contrasts with other machaeroidines in having a high Zx / Zy canine value (2.16; Fig. 5B View Figure 5 ). Such a value, although within the range of machairodonts ( Figs 9 View Figure 9 , 10), is still low compared to other flanged sabretooths ( Figs 6, 7 View Figure 7 , 8 View Figure 8 , 11 View Figure 11 ). Nevertheless, it does indicate that stresses were better constrained dorsoventrally in the symphysis when M. simpsoni delivered its canine bite than in other machaeroidines. Thus, M. simpsoni may have been able to immobilize its prey more efficiently than other machaeroidines prior to delivering canine bites.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.