
SPARASSODONTA
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00194.x |
|
persistent identifier |
https://treatment.plazi.org/id/CB5187EB-FFAD-FF94-4F95-FAA2BAF6A670 |
|
treatment provided by |
Diego |
|
scientific name |
SPARASSODONTA |
| status |
|
THYLACOSMILINAE, SPARASSODONTA
Represented by a single, long-lived species, Thylacosmilus atrox , this clade of dirk-toothed marsupials lived in South America from the late Miocene until the late Pliocene ( Fig. 1 View Figure 1 ) ( Pascual & Bondesio, 1982; Goin & Pascual, 1987). Extensive work has been done on the craniodental and postcranial adaptations of thylacosmilines and their comparison with those of eutherian sabretooths ( Riggs, 1934; Marshall, 1976; Turnbull, 1976, 1978; Churcher, 1985; Goin & Pascual, 1987). The different dental formula (I 1, C 1, P 2, M 4) and absence of a carnassial tooth homologous to that of carnivorans in Thylacosmilus (the fourth molar is modified into a shearing blade) requires additional measurement sites. Even though the thylacosmiline interdental gaps may not be functionally equivalent to those of eutherians, they give an overall appreciation of the force variation along the mandible (from post- M 4 to P 2 P 3, and behind the canine). Both an adult (P14531, holotype) and a juvenile individual (P14344, paratype) were studied in order to elucidate differences in mandibular force profiles, which could give insight into changes in feeding behaviour occurring during thylacosmiline ontogeny.
A brief note on the proper orientation of the Thylacosmilus mandible must be made. Originally, Riggs (1934: plate 3) believed the sabres of Thylacosmilus to be strongly divergent. Consequently, he reconstructed the mandible with laterally flaring mandibular flanges, even though this resulted in a shorter mandibular symphysis than indicated by the rugosity on the medial aspect of the flanges. Later studies revealed that the sabres were not divergent and that the mandibular symphysis extended nearly to the lower extremity of the mandibular flange ( Turnbull, 1976; Churcher, 1985; Goin & Pascual, 1987), which resulted in a slight reorientation of the mandibular corpus. Beam models designed for both mandibular orientations have delivered nearly identical results, except for the values at the canine due to the shorter mandibular symphysis of Riggs’ (1934) reconstruction. Owing to the similarity of results, only the model for the revised mandibular orientation will be discussed.
Mandibular force profiles
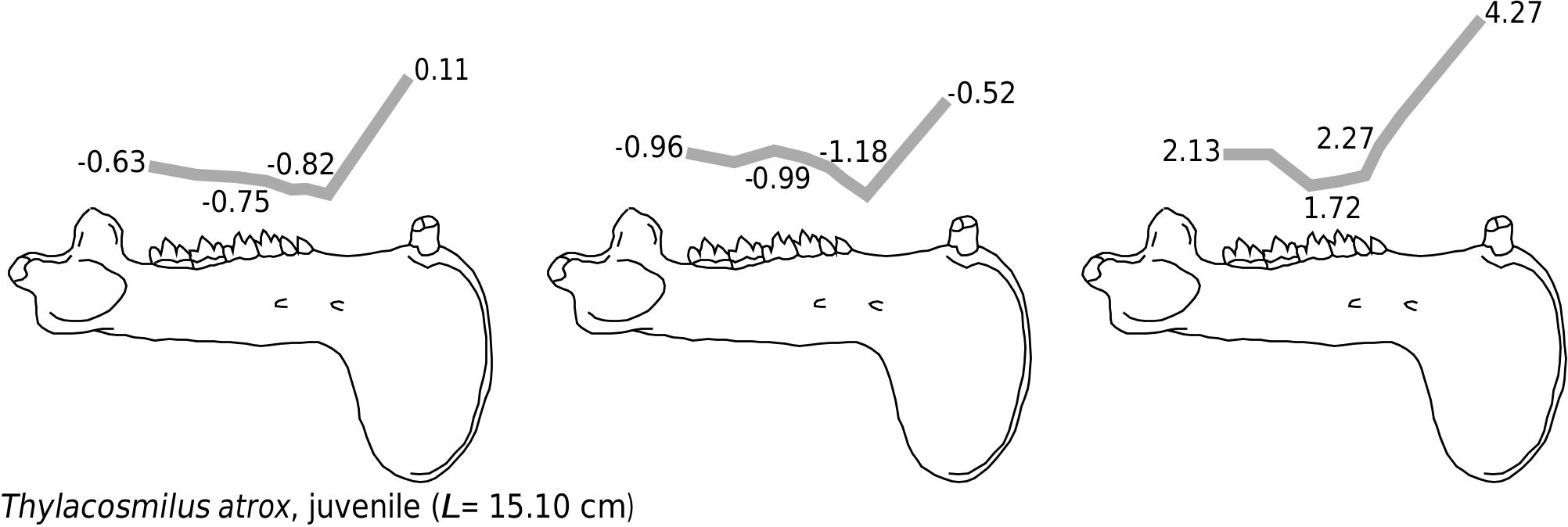
Other than differing in absolute values, the profiles of the two thylacosmilines are nearly identical ( Fig. 11 View Figure 11 ). Dorsoventrally (Zx / L), mandibular force values decrease slowly along the cheek teeth, attaining a minimum at P 2 P 3. Labiolingually (Zy / L), the mandibular force values remain relatively constant over the molars and decrease rapidly over the premolars, attaining a minimum at P 2 P 3. The mandible at the canine is extremely strong, both dorsoventrally and labiolingually, being much stronger than at the carnassial-analogue M 4.
Juvenile and adult Thylacosmilus differ the most from other carnivorous mammals and, surprisingly, from each other in their relative mandibular force profile (Zx / Zy; Fig. 11 View Figure 11 ). In the adult, the relative mandibular force profile remains relatively constant (Zx / Zy = 1.50) over the entire tooth row, the mandible being slightly narrower behind the last molar (Zx / Zy = 1.80). These values are lower than those observed in carnivorous eutherians. In the juvenile, the mandible maintains a relatively constant shape under the anteriormost molars (Zx / Zy ≈ 1.72), but is narrower under the last molar and the premolars (Zx / Zy> 2.13). Finally, the symphyseal region is extremely deep in thylacosmilines, with Zx / Zy canine values rivalling those of Barbourofelis fricki and Hoplophoneus dakotensis . The juvenile Thylacosmilus has higher Zx / Zy canine values than the adult (4.27 and 3.15, respectively), although it is impossible to evaluate the statistical significance of this difference.
Interpretation
In addition to assuming similarity of cortical bone thickness and of safety factors for bite force estimation purposes, tooth homology must also be considered when comparing eutherians and metatherians. Because the M 3 M 4 interdental gap in Thylacosmilus is situated at the same relative position as the P 4 M 1 interdental gap in sabretoothed eutherians (∼42–44% of mandibular length), although slightly farther back than in conical-toothed carnivorous eutherians and marsupials (∼50%; Emerson & Radinsky, 1980; Goin & Pascual, 1987; Werdelin, 1987; Bryant & Russell, 1995), comparison with the eutherian precarnassial interdental gap (P 4 M 1) is considered valid. The biomechanical approach indicates that Thylacosmilus could exert a bite force 34% greater than Panthera tigris , a felid of similar mandibular length ( Table 1).
Both the dorsoventral and labiolingual force profiles show that the symphyseal region of Thylacosmilus was stronger in bending than the mandibular corpus near the cheek teeth, indicating that this predator delivered powerful canine bites. The very shallow negative slope formed by the Zx / L -values along the mandibular ramus suggests that force was uniformly applied along the entire tooth row ( Fig. 11 View Figure 11 ). In order to explain the unusual labiolingual force profile, one must look at the shape of the mandibular ramus. As was the case in barbourofelids and smilodontines, the mandibular ramus is labially inclined near the cheek teeth, being at its farthest lateral position at M 2 (also see Riggs, 1934); this feature is even more obvious in the juvenile specimen studied. The labial rotation of the mandibular ramus slightly increases the labiolingual diameter near the molars, explaining the high labiolingual force over the posterior molars.
The relatively constant Zx / Zy values between M 4 and P 2 in the adult thylacosmiline ( Fig. 11 View Figure 11 ) reflect the uniform shape of the mandibular ramus (see Riggs, 1934; Churcher, 1985; Goin & Pascual, 1987). The constancy of the Zx / Zy ratio indicates that the ramus is not adapted to sustain localized high loads, such as in bone cracking, but for the unique purpose of slicing flesh (Therrien, 2005). This interpretation is corroborated by the dentition of thylacosmilines, which is unique among carnivorous marsupials in being specialized for pure slicing ( Riggs, 1934; Churcher, 1985; Goin & Pascual, 1987). Even the highly carnivorous borhyaenines, the sister taxon to thylacosmilines, possessed robust premolars that have been interpreted as an adaptation for bone cracking ( Marshall, 1978). The lower Zx / Zy values of the mandibular ramus in the adult thylacosmiline relative to other carnivorous
Dorsoventral force Labiolingual force Relative force (Log Zx/L) (Log Zy/L) (Zx/Zy)
Thylacosmilus atrox , adult (L = 20.54 cm)
mammals is presumably an artefact of the lateral rotation of the mandibular corpus. The fact that the lowest Zx / Zy values in the juvenile thylacosmiline correspond to the farthest lateral extension of the tooth row strongly supports this interpretation.
The high Zx / Zy canine values of thylacosmilines indicate that the symphyseal region was extremely well buttressed dorsoventrally and that loads were well constrained within the sagittal plane during the canine bite ( Fig. 11 View Figure 11 ). Similar buttressing is achieved only by the dirk-toothed predators H. dakotensis and B. fricki . Consequently, Thylacosmilus must have used its sabres against a restrained or immobilized prey, for struggling prey would have induced unpredictable stresses for which the mandible was not designed to sustain. The reduced incisor battery of Thylacosmilus argues against any significant grasping function for the anterior extremity of the mandible ( Goin & Pascual, 1987) but its appendicular skeleton best resembles that of eutherian dirk-toothed sabretooths. The humerus is extremely robust with a well-developed deltopectoral crest, indicative of a powerful forelimb ( Riggs, 1934; Argot, 2004a, b). The humeral and ulnar articulations of the radius, combined with the prominent lateral epicondylar crest of the humerus, indicate that the forelimb was capable of extensive pronation/ supination movements for grasping ( Riggs, 1934; Argot, 2004b). The presence of a semiopposable pollex in Thylacosmilus further supports the interpretation that the forelimbs were used for grasping ( Argot, 2004a, b). Although the radius is incomplete, Riggs (1934: 25) stated that it ‘appears to have been almost as large as the tibia’; one must presume that Riggs referred to the tibia of the paratype (juvenile) because the holotype (adult) lacks this element. If this is the case, then the brachial index for the adult Thylacosmilus would have been near 0.75 (derived from measurements in Riggs, 1934), lower than Smilodon fatalis but similar to Barbourofelis loveorum (see above; Baskin, in press). The hindlimb was also robust. The femur has a straight shaft and a prominent, rounded head, a condition unlike that of any other marsupial but reminiscent of bears and eutherian sabretooths ( Riggs, 1934; Argot, 2004a, b). Although a complete tibia is not preserved in the adult, the crural index of the juvenile Thylacosmilus , derived from dimensions reported by Riggs (1934), is 0.71, which is also slightly lower than S. fatalis but within the range of B. loveorum and extant bears (see above; Anyonge, 1996; Baskin, in press). These characteristics do support the hypothesis that Thylacosmilus , like dirk-toothed sabretooths, was a powerful ambush predator capable of pulling down large prey and immobilizing it with its forelimbs (see Argot, 2004a, b).
Despite the clearly robust postcranial morphology of Thylacosmilus, Goin & Pascual (1987) argued that this predator could not have subdued large prey (e.g. macraucheniids, megatheriids, mylodontids or toxodonts) with its forelimbs because: (1) these prey would have been too large to be tackled by the ‘cougar-size’ Thylacosmilus ; (2) the absence of gregarious hunting behaviour among living polyprotodonts argues against collaborative hunting in Thylacosmilus to subdue large prey; and (3) thylacosmilines did not possess retractile claws, which would have allowed them to grasp prey. Instead, the authors suggested that Thylacosmilus rammed its prey in order to knock it down, its robust head being capable of absorbing the shock of the impact, a highly unusual, if not improbable, technique.
Two of the three arguments employed by Goin & Pascual (1987) to discredit a dirktooth-like behaviour in thylacosmilines are invalid. First, the holotype of Thylacosmilus (P14531) has cranial and mandibular dimensions comparable to those of a tiger (the skull being only slightly shorter due to the vertically orientated occipital region), thus suggesting an animal much larger than a cougar ( Puma concolor ). Additionally, the robust appendicular skeleton of the adult Thylacosmilus , almost bear-like in proportions [compare intermembral indices presented above with Anyonge’s (1996) results], suggests that it could have tackled large prey (if not fully-grown adults of the aforementioned prey species, then at least juveniles and/or subadults). Secondly, other than among canids and hyaenids, gregariousness is uncommon among eutherian carnivores; even among modern felids feeding on large prey, collaborative hunting is the exception rather than the rule ( Schaller, 1972; Scheel & Packer, 1991; Caro, 1994). Gregarious behaviour among eutherian sabretooths has neither been clearly established (see McCall, Naples & Martin, 2003), nor is it a requisite for their inferred killing technique on large prey. Recently, Van Valkenburgh & Sacco (2002) investigated sexual dimorphism in Rancho La Brea carnivores and observed that the degree of sexual dimorphism exhibited by S. fatalis suggested that this animal lived either in a typical solitary felid social organization or in a monogamous breeding system like that of wolves (with one dominant pair and offspring from the current and/or previous years), but clearly did not form prides like African lions.
Thus, the dramatic ramming behaviour inferred by Goin & Pascual (1987) rests solely on the apparent lack of retractile claws in Thylacosmilus , as inferred on the basis of a single proximal half of an ungual bearing likeness to those of Borhyaena tuberata (see Riggs, 1934) and the absence of such features among marsupials in general. Re-examination of the type material of Thylacosmilus by Argot (2004b) revealed that none of the osteological correlates of claw retraction (see Gonyea & Ashworth, 1975) are present. It is conceivable, albeit just a supposition, that the ever-growing sabres of Thylacosmilus could have evolved in response to a reduced efficiency to immobilize prey. In strong contrast with eutherian sabretooths, Thylacosmilus possessed a reduced incisor battery and lacked retractile claws, which raise the possibility that this predator would not have been as good at subduing prey as its eutherian analogues. Consequently, the risk of accidental sabre breakage would have been increased, particularly during the initial phase of the attack when torsional stresses are at their greatest. Ever-growing sabres may have been the solution adopted by thylacosmilines to compensate for their less efficient grappling adaptations due to their borhyaenoid phylogenetic heritage (e.g. Argot, 2003, 2004a, b).
Finally, the higher Zx / Zy canine values of juvenile Thylacosmilus relative to the adult is reminiscent of the situation observed in modern felids characterized by extended periods of parental care (Therrien, 2005). Although caution is advisable given the limited sample size, this interpretation is not unreasonable. Indeed, the study of dental eruption sequence in Homotherium serum ( Rawn-Schatzinger, 1983) , S. fatalis ( Tejada-Flores & Shaw, 1984) , and Barbourofelis species ( Bryant, 1988, 1990) has revealed that juvenile eutherian sabretooths underwent extended parental care and became independent around the same age as modern felids. During this extended period of parental care, juvenile sabretooths would have learned and honed their skills to kill prey with sabre-like canines under adult supervision while they fed on prey killed by a parent.
In juvenile S. fatalis , the deciduous sabre (dC 1) is preserved until the permanent canine attains a similar length (c. 65–70 mm). At that time, dC 1 is shed and the permanent sabre pursues its eruption. Therefore, juveniles do not undergo a stage where they lack functional sabres ( Tejada-Flores & Shaw, 1984). Comparison of microwear on deciduous and permanent upper canines revealed that juveniles were not as precise or careful with their sabres as adults ( Akersten, 1985), possibly reflecting a learning period. Consequently, juveniles could have practised their killing technique with their deciduous canines. By the time the permanent sabres were in place, the individual had refined its killing techniques and would have been apt to use the sabres efficiently and safely.
In contrast, the dC 1 in Barbourofelis , and most nimravids, is nearly as large as the permanent sabre (from 80% to 98% its length) and its eruption is delayed until the permanent cheek teeth have erupted, an equivalent stage of dental eruption to the permanent canines of extant lions ( Bryant, 1988, 1990). Thus, juvenile barbourofelids underwent a long ontogenetic interval without functional upper canines before the large dC 1 started acting as ‘a second pair of permanent sabres at the expense of the deciduous pair and hence a fully functional replacement in the event of breakage to the first pair early in life’ ( Bryant, 1988: 305).
With its ever-growing upper canines, Thylacosmilus probably did not suffer too seriously from the detrimental consequences of sabre breakage. Juveniles could have learned gradually how to use their sabres and in the event that they broke a sabre accidentally, owing to inexperience and/or carelessness, the tooth would have eventually regrown to its original size. The sabre could then have been resharpened through thegosis against the lower canines ( Goin & Pascual, 1987).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.