
Ronzotherium velaunum
|
publication ID |
https://doi.org/10.5852/ejt.2021.753.1389 |
|
publication LSID |
lsid:zoobank.org:pub:8009DD3B-53B0-45C9-921E-58D04C9C0B48 |
|
DOI |
https://doi.org/10.5281/zenodo.4959435 |
|
persistent identifier |
https://treatment.plazi.org/id/C53FFD4D-D70D-FFB5-FE7F-DF6B1B74A6DB |
|
treatment provided by |
Felipe (2021-06-14 14:00:33, last updated 2024-11-27 04:33:04) |
|
scientific name |
Ronzotherium velaunum |
| status |
|
Ronzotherium velaunum (Aymard in Pictet, 1853)
Figs 2–7 View Fig View Fig View Fig View Fig View Fig View Fig
Acerotherium velaunum Aymard in Pictet, 1853: 296.
Ronzotherium cuvieri Aymard, 1856: 233 .
Rhinoceros velaunus – Aymard in Pictet 1853: 298.
Rhinocéros View in CoL à incisives ( Ronzotherium ) – Aymard 1854: 675.
Ronzotherium velaunum – Aymard 1856: 233. — Filhol 1881: 3. — Osborn 1900: 232–237, 241, fig. 3. — Deninger 1903: 94–95. — Stehlin 1909: 509. — Abel 1910: 4–6, 8–9, 18, 33. — Roman 1912a: 4–5, 8, 10. — Kafka 1913: 5, 47, fig. 40a. — Airaghi1925: 25. — Heissig 1969: figs 6a, 8c, 9a, 10a, 11, 25a (from Ronzon). — Brunet 1977: 16, 23; 1979: 102–104, 152–153, table 51, pls XV, XIXa–f. — Brunet et al. 1977: 109–112. — Jehenne & Brunet 1992: 202–203. — Uhlig 1996: 140–142. — Ménouret & Guérin 2009: 293–327. — Becker 2009: 495, 500.
Ronzotherium cuvieri (?) – Filhol 1881: 3.
Acerotherium velaunum – Filhol 1881: 75–78, figs 69–86, 88. — Mermier 1895: 176, 180, 186. — Roman 1910: 1558–1560; 1912a: 7, 27, 42–45, 56, 78, fig. 13, pl. II figs 2, 2a. — Gignoux 1928: 147, 149, 151.
Acerotherium cuvieri – Filhol 1881: fig. 87, 89–90. — Airaghi 1925: 26, 29.
? Ronzotherium cf. velaunum – Schlossser 1902: 112–113, pl. V figs 23, 25.
Rhinoceros velaunus – Roman 1912a: 45.
? Ronzotherium velaunum – Kafka 1913: 48–50, figs 40b, 41. — Kretzoi 1940: 89–92, 97–98, figs 1–2. — Lavocat 1951: 115. — Balme 2000: 153. — Costeur & Guérin 2001: 77.
Rhinoceros velaunum – Airaghi 1925: 32–33, 40–41.
Ronzotherium cf. velaunum – Heissig 1978: 249.
Non Ronzotherium filholi – Lavocat 1951: 116, pl. 19 fig. 3, pl. 26 fig. 1 (from Vendèze).
Non Ronzotherium velaunum – Heissig 1969: figs 5, 6b–d, 7, 8a–b, d–g, 9b–c, 10b–d, 25b (from ‘Auvergne’, Mouillac, Vendèze, St-Henri, St-André, Marseille, Les Milles).
Historical diagnoses
The first diagnosis of the species was provided by Heissig (1969, translated by the authors): “ type species of the genus Ronzotherium with almost parallel i2 facing forward; i1 absent, I1 and I2 large. Lower jaw branches at an acute angle to each other. Upper molars broad, with long postfossette, narrow, slightly curved medisinus, thick and far forward paracone and mostly weak or missing lingual cingulum; M3 with sharp, narrow ectoloph edge behind the metacone. Upper premolars with straight or barely curved, parallel, originally slightly inclined transverse lophs and strongly waved lingual cingulum, slowly reduced; reduction begins at P4. P2 semimolariform to molariform, P3 and P4 premolariform to submolariform, but with relatively far apart inner lophs. Lower molars broad with weak labial cingulum; lower premolars with long talonid, mostly groove-shaped talonid pit and sharp, deep external groove. The entoconid lies far back, the cingulum is weak. The p1 is single rooted or missing.”
An emended diagnosis was provided by Brunet (1979, translated by the authors): “Stratigraphically the most ancient and primitive species of its kind. Skull: unknown. Mandible: posterior border of the symphysis just ahead of the d1, its lower surface presents a hull; very strong occlusion between i1 and i2. Decidual teeth: the upper milk premolars are unknown; the inferiors have a strongly curved hypolophid; d1 is biradiculate; the first lobe of d2 is strong with a long lingual branch of the paralophid, the ‘metaconid’ is not individualized; the anterior lobe of d3 is strong with a very long anterior branch of the paralophid. Definitive dentition: probable presence of i1. Upper premolars with a short postfossette, located above the posterior cingulum; strong lingual cingulum, barely waved. Upper molars with strong lingual cingulum, complete or disappearing only at the level of the hypocone. Lower premolars and molars: more or less large with a strong labial cingulum, more or less complete; the very notched talonid fossae on the labial side of the hypolophid are flatter, more horizontal, and lingually higher than in R. filholi ; the trigonid fossae also open higher, above the anterolingual cingulum; premolars with long paralophid, without protoconid fold; P2 not reduced, with a strong anterolabial groove. Appendicular skeleton: tetradactyl hand with a gracile McV, reduced but complete; on the dorsal side of the hand, the lunate articulates with the magnum; on the pyramidal, the ulnar facet is more laterally widened and the lower facet for the lunate higher and larger than in R. filholi ; likewise, the magnum carries a much longer and higher facet for the McII.”
Emended diagnosis
Type species of the genus with a posterior border of the symphysis located anterior to p2 and without lingual groove for the sulcus mylohyoideus on the corpus mandibulae. The metacone fold is present on M1–2. The d1 is absent in the juvenile, and the entoconid is constricted on decidual lower milk teeth. The cingula are poorly developed on upper and lower cheek teeth and discontinuous. The posteroproximal and anteroproximal facets for the lunate are in contact on the scaphoid and the fibula facet is oblique on the astragalus. The trapezium facet is absent on the McII.
Type material
Lectotype
FRANCE • right hemimandible still partly in sediment with poorly preserved p2–m3 and broken symphysis; Haute-Loire, near Le Puy-en-Velay, hill of Ronzon ; PUY.2004.6.1765.RON .
Additional material
FRANCE • 1 broken mandible in several pieces, with i2 and p2–m3 on the left side and i2 and p2–(m1) on the right side; same collection data as for lectotype; PUY.2004.6.1766.RON • 1 juvenile mandible, still partly in sediment, with d2–d4 and erupting m1 on both sides and a small di1; same collection data as for lectotype; PUY.2004.7.1.RON • 1 broken ectoloph of P2?; same collection data as for lectotype; PUY.2004.6.1551.RON • 1 isolated P3; same collection data as for lectotype; PUY.2004.6.1767.RON • 1 isolated M1; same collection data as for lectotype; TLM.PAL.2010.0.122 • 1 cast of an isolated lower molar; same collection data as for lectotype; PUY.2004.6.841.RON • 1 distal part of humerus; same collection data as for lectotype; PUY.2004.6.262.RON • 1 complete scaphoid; same collection data as for lectotype; MNHN.F.RZN.503 • 1 lunate partly unextracted from sediment; same collection data as for lectotype; PUY.2004.6.1901.RON • 1 pyramidal, still in sediment; same collection data as for lectotype; MNHN.F.RZN.504 • 1 pyramidal; same collection data as for lectotype; MNHN.F.RZN.502 • 1 pisiforms, still in sediment; same collection data as for lectotype; MNHN.F.RZN.505 • 1 pisiforms, still in sediment; same collection data as for lectotype; PUY.2004.6.1901.RON • 1 magnum, still in sediment; same collection data as for lectotype; PUY.2004.6.907.RON • 1 magnum; same collection data as for lectotype; PUY.2004.6.263.RON • 1 broken anterior part of unciform; same collection data as for lectotype; PUY.2004.6.1480.RON • 2 distal parts of femora; same collection data as for lectotype; PUY.2004.6.266.RON , PUY.2004.6.267.RON • 2 proximal parts of tibiae; same collection data as for lectotype; PUY.2004.6.260.RON , PUY.2004.6.261.RON • 1 ectocuneiform, still partly in sediment; same collection data as for lectotype; PUY.2004.6.577.RON • 1 cuboid, still in sediment; same collection data as for lectotype; PUY.2004.6.1309.RON • 1 cuboid; same collection data as for lectotype; PUY.2004.6.268.RON • 1 astragalus, still preserved in sediment; same collection data as for lectotype; PUY.2004.6.1770.RON • 1 central metapodial, still in sediment; same collection data as for lectotype; PUY.2004.6.840.RON • 1 lateral phalanx, still in sediment; same collection data as for lectotype; PUY.2004.6.604.RON .
Type horizon and locality
Hill of Ronzon, near Le Puy-en-Velay (Haute-Loire, France), MP21 (early Oligocene).
Stratigraphical distribution
MP21 (early Oligocene).
Geographical distribution
France: Ronzon, Lagny-Torigny, Ruch. Germany: Haag 2, Möhren 20.
Description
MANDIBLES. Three mandibles of R. velaunum from Ronzon are preserved. The lectotype mandible PUY.2004.6.1765.RON is a right hemimandible with p2–m3 ( Fig. 2A–D View Fig ). The posterior part of the specimen and the symphysis are broken, and the left side is still in sediment. The base of the corpus mandibulae is straight and low, with a constant height below the teeth neck. The ramus is vertical, and the coronoid process is well developed and high. The mandible PUY.2004.6.1766.RON is badly preserved and in several pieces ( Fig. 2E–J View Fig ). The symphysis as well as both branches are preserved, with i2, the root of d1 and p2–m3 on the left side, and only i2 and p2–m1 on the right side. It was recently prepared and new characters can now be observed: the angle between the symphysis and the corpus is low, the symphysis is rather narrow and its posterior borders is in front of p2, the foramen mentale is below p2 and there is no lingual groove of the sulcus mylohyoideus. The last mandible PUY. 2004.7.1.RON belonged to a juvenile individual and is still partly preserved in sediment ( Fig. 2K–O View Fig ). It bears d2–d4 and erupting m1 on both sides as well as a small di1 on the right side. There is apparently no dp1. The posterior border of the symphysis is anterior to d2. No lingual groove of the sulcus mylohyoideus is visible.
UPPER DENTITION. Very few upper teeth are preserved in this locality ( Fig. 3 View Fig ): an ectoloph of a left P2 (PUY.2004.6.1551.RON), a P3 (PUY.2004.6.1767.RON) and an M1 (TLM.PAL.2010.0.122). However, Filhol (1881) noted the existence of an upper maxilla that he could not have accessed during his study and was supposedly in Pichot-Dumazel’s collection. Unfortunately, this maxilla remains unknown. The P2 and P3 have strong paracone and metacone folds and very thin discontinuous labial cingulum. Their crown is low. The lingual cingulum is strong and continuous on P3. The P3 is three-rooted and few characters can be observed, as it is very worn. Its postfossette is narrow and the protocone and hypocone were probably not separated. The M1 has four roots and is also much worn. Labial cingulum is almost completely absent. Lingual cingulum is strong and continuous under the protocone and disappears under the hypocone. The paracone fold is strong and the metacone fold is present but very thin. The parastyle is strong and there is no mesostyle. The protocone does not seem constricted. The posterior profile of the ectoloph is slightly concave.
LOWER DENTITION. The definitive anterior dentition is only represented by two i2 from the mandible PUY.2004.6.1766.RON. They are straight and horizontal. The roots are wider than the crown, and the crown shows a clear and large wear-facet, which means that I1 and i2 could contact each other. The transverse outline of the crown is in the shape of a medially pinched drop. The neck is not marked and the enamel is very thin. The lower cheek teeth are two-rooted and low-crowned. There is no cement. The premolar row is short compared to the molar row (0.42 <Lp3–4/Lm1–3 <0.50). A weak labial cingulum is sometimes present on the lower cheek teeth, but a lingual cingulum is always absent. Vertical external rugosities are present on the ectolophid of p2–3. The ectolophid groove is developed and does not vanish before the neck. In occlusal view, the trigonid is very angular and forms a right dihedron which becomes more acute with wear, while the talonid is rounded. The talonid basin of the lower premolars is poorly developed: the entoconid is completely absent and the hypoconid is low. The hypolophid vanishes before the posterolingual border of the premolars, the posterior valley is therefore very wide and U-shaped. On the contrary, the anterior valley is narrow, and both valleys open very high above the neck. The metaconid is the largest and most developed cusp on lower premolars. On p3, the metaconid bears an anterior crest, almost closing the anterior valley. The paralophid of premolars has two branches, a labial branch, and a high and long anterior branch, parallel to the protolophid. The molars greatly differ from the premolars by the much stronger development of the entoconid, which is also slightly constricted. The opening of the anterior valley is higher than the posterior one.
DECIDUAL DENTITION. Only the lower decidual dentition is known from Ronzon, from the juvenile mandible PUY. 2004.7.1.RON ( Fig. 2K–O View Fig ). The di1 is very small and has a conical crown. There does not seem to be a d 1 in the juveniles. However, d2–4 are well developed. The metaconid and entoconid are slightly constricted, especially on d4. There is neither a protoconid fold nor a vertical external rugosity. The lingual and labial cingulum are absent. The ectolophid fold is strong on d2 but there is no anterior groove on the ectolophid. The paralophid is double on d2–3 and simple on d4. On d2, the posterior valley is almost closed by the extension of the entoconid, but still narrowly open. There is no lingual groove of the entoconid on d3. The d4 is very molariform.
HUMERUS. One distal fragment of humerus is preserved (PUY.2004.6.262.RON, Fig. 4A–C View Fig ). The fossa olecrani is high but not very deep. The distal articulation is well constricted and there is no scar on the trochlea. The distal gutter on the epicondyle is also absent. Medial and lateral epicondyles are poorly developed and the lateral epicondylar crest is weakly extended laterally.
SCAPHOID. The scaphoid MNHN.F.RZN.503 ( Fig. 4D–G View Fig ) is well preserved. The anterior height is equal to the posterior one. The postero-proximal articulation with the lunate bone is not visible but may have been present on the eroded proximo-lateral tuberosity and fused with the anteroproximal facet. The proximal facet for the radius is very concave and fuses anteriorly with the anteroproximal facet for the lunate bone. The anterodistal facet for the lunate is poorly distinguished. The three distal articular facets are concave in lateral view. The trapezium facet is rather large and triangular. The trapezoid facet is the largest and has a prominent dorso-medial extension. The magnum facet is concave in lateral view.
LUNATE. The lunate bone PUY.2004.6.1901.RON ( Fig. 4H–J View Fig ) is still mostly concealed in the sedimentary block. Only the proximal, dorsal and lateral sides are visible. It is an overall large and robust bone. The posterior tuberosity is almost as wide as the proximal facet for the radius. Two articular facets are visible on the lateral side, both corresponding to the pyramidal bone. The proximal facet is small while the distal one is large, flat and circular. On the medial side, two well separated articular facets can be distinguished and correspond to the scaphoid, which implies the presence of a postero-proximal facet for the lunate on the scaphoid, that is not visible on the scaphoid MNHN.F.RZN.503.
PYRAMIDAL. Two pyramidals are preserved (MNHN.F.RZN.502, Fig. 4K–M View Fig and MNHN.RZN.504, Fig. 4N View Fig ). There are two proximal articulation facets: a large one for the ulna, and a smaller one, elongated and band-like for the pisiform. The medio-distal articulation for the lunate is symmetrical and the distal facet for the unciform is triangular.
PISIFORM. The pisiform MNHN.RZN.505 is still in articulation with the pyramidal MNHN.RZN.504. Another unnumbered pisiform is preserved on the sedimentary bloc of the lunate bone PUY.2004.6.1901. RON. The pisiform is very small, and neither flattened nor elongated. It bears a large proximal articular facet for the radius. The distal end is roughly conical and rounded.
UNCIFORM. Only the dorsal part of the left unciform PUY.2004.6.1480.RON is preserved, the posterior tuberosity is missing ( Fig. 5F–G View Fig ). There are two proximal facets: a large one, dorso-ventrally convex for the pyramidal, and smaller one, flattened and arrowhead-shaped for the lunate. They form an angle of 120–130° in dorsal view. The posterior expansion of the pyramidal facet is very short and wide. The three distal facets, for the magnum, McIII and McIV, are partially covered in sediment. The lateral McV facet is broken but was probably distinct from the pyramidal facet.
MAGNUM. Two magnums are preserved. PUY.2004.6.907.RON is still in a sedimentary bloc, while PUY.2004.6.263.RON is subcomplete and fully extracted ( Fig. 5A–E View Fig ). It is a rather tall bone, the proximodistal height is almost equal to the dorsoventral length, but it is very compressed transversally. In anterior view, the anterior border of the scaphoid facet is nearly straight. The lunate facet is very long dorsoventrally, and very convex proximally. There are two medial facets below the scaphoid facet: a proximal one for the trapezoid and a distal one for the McII. The former is trapezoidal while the latter is curved. There is no indentation between these two facets. The distal facet for the McIII is large and deeply concave dorsoventrally. The unciform facet on the lateral side is not preserved. The posterior tuberosity of the magnum is long, thin and curved.
FEMUR. There are two distal ends of left femora in Ronzon (PUY.2004.6.266.RON, Fig. 6A–D, I View Fig and PUY.2004.6.267.RON, Fig. 6E–H, J View Fig ). In anterior view, the medial lip of the trochlea is prominent. The groove between the two trochlea is not very deep and the proximal border of the trochlea is almost straight. In lateral view, the medial lip of the trochlea is strongly forward compared to the diaphysis. In posterior view, the two condyles are similar in size and widely separated by the intercondylar fossa.
The supracondylar fossa is shallow. In distal view, the articular surfaces of the trochlea and the condyles are connected medially and laterally.
TIBIA. Two proximal ends of left tibias (PUY.2004.6.260.RON and PUY.2004.6.261.RON) could belong to the same individuals as the femora ( Fig. 6K–P View Fig ). In proximal view, it is wider than long. The tibial tuberosity is weakly developed and is laterally displaced. It is separated from the medial tuberosity by a wide groove. The cranial intercondylar area is deep and wide, the central one very small and the caudal one is deep and slender. The lateral condyle is oval, and wider than long, while the medial one is almost rectangular and longer than wide. In anterior view, the medial tuberosity is higher than the lateral one. In lateral view, the groove for the extensor is wide and shallow and the tibial fossa rather deep. The tibia and fibula were completely independent, there is no contact mark along the diaphysis, only a high articular facet below the lateral condyle.
ASTRAGALUS. Only the anterior face of the astragalus (PUY.2004.6.1770.RON) is visible, the other side is still in sediment, but it is complete ( Fig. 7A–D View Fig ). The transverse diameter/height (TD/H) ratio is slightly above 1, but below 1.2, whereas the anteroposterior diameter/height (APD/H) ratio is below 0.65. On the lateral side, the fibula facet is slightly oblique and flat. The collum tali is very high. There are two distal articular facets: the navicular facet is large and slightly concave transversally, while the facet for the cuboid is small and flat. In distal view, the trochlea is very oblique compared to the distal articulation. The medio distal tubercle is well developed.
CUBOID. Two cuboids are preserved: one is still partially in sediment (PUY.2004.6.1309.RON) but the other is subcomplete (PUY.2004.6.268.RON, Fig. 7I–M View Fig ). The proximal articular surface is triangular. There are two distinct surfaces, for the astragalus and the calcaneus, distinguished by a shallow groove. The calcanear one is the largest. In anterior view, the bone is rectangular and higher than wide. In lateral view, the lateral groove for the tendons is very deep. The posterior apophysis is wide and stout, and extends more distally than the distal articular facet. The distal articulation surface for the MtIV is almost a right triangle with rounded edges.
ECTOCUNEIFORM. The right ectocuneiform PUY.2004.6.577.RON is still partially in sediment, the proximal side is not visible ( Fig. 7E–H View Fig ). The distal articular facet for the MtIII is crescent-shaped. The posterolateral process is rather short and medially oriented. The medial side is straight and bears three facets: one dorsal and band-shaped for the mesocuneiform, and two distal, oval-shaped for the MtII. The lateral side is curved and the two articulations postero-proximal and anterodistal for the cuboid are separated by a deep groove.
METAPODIAL. A central metapodial (PUY.2004.6.840.RON) is also preserved from Ronzon, still in sediment, and only the dorsal side is visible ( Fig. 5H–I View Fig ). The proximal articulation is very incomplete, but it is nonetheless rather dorsoventrally flat, which would indicate a MtIII rather than a McIII, as also suggested by Brunet (1979). There is a small anteroproximal facet for the MtII, the posterior one, if present is hidden by sediment. The diaphysis gets slightly wider towards the distal end. The median keel of the distal articulation is smooth.
LOST MATERIAL. The scaphoid and pyramidal thought as lost by Brunet(1979) and figured by Filhol (1881) are now in fact in the collections of MNHN (Paris, France) (MNHN.F.RZN.502, MNHN.F.RZN.503 and MNHN.F.RZN.504). However, the calcaneum, MtIV and McV, figured by Filhol (1881: pl. 13), are indeed lost and could not be found either in the Musée Crozatier (Le Puy-en-Velay, France) or in the MNHN.
Abel O. 1910. Kritische Untersuchungen uber die palaogenen Rhinocerotiden Europas. Abhandlungen der geologischen Reichsanstalt, Wien 20 (3): 1 - 52.
Airaghi C. 1925. Considerazioni filogenetiche sui Rinoceronti d'Europa. Rivista Italiana di Paleontologia 31: 23 - 46.
Aymard A. 1854. Des terrains fossiliferes du bassin superieur de la Loire. Comptes rendus des Seances de l'Academie des Sciences, Paris 38: 673 - 677.
Aymard A. 1856. Rapport sur les collections de M. Pichot-Dumazel. Congres scientifique de France 22: 227 - 245.
Balme C. 2000. Decouverte d'empreintes de pas de mammiferes fossilisees dans la Carriere d'argile de Triclavel, Commune de Viens (Vaucluse). Courrier scientifique du Parc naturel regional du Luberon 4: 152 - 155.
Becker D. 2009. Earliest record of rhinocerotoids (Mammalia: Perissodactyla) from Switzerland: systematics and biostratigraphy. Swiss Journal of Geosciences 102 (3): 489 - 504. https: // doi. org / 10.1007 / s 00015 - 009 - 1330 - 4
Brunet M., Jehenne Y. & Ringeade M. 1977. Note preliminaire concernant la decouverte d'une faune et d'une flore du niveau de Ronzon dans l'Oligocene inferieur du Bassin d'Aquitaine. Geobios 10 (1): 109 - 112. https: // doi. org / 10.1016 / S 0016 - 6995 (77) 80057 - 0
Brunet M. 1979. Les grands mammiferes chefs de file de l'immigration Oligocene et le probleme de la limite Eocene - Oligocene en Europe. Fondation Singer-Polignac, Paris.
Costeur L. & Guerin C. 2001. Les pistes et empreintes de mammiferes de l'Oligocene ancien de Viens (Vaucluse). Courrier scientifique du Parc naturel regional du Luberon 5: 74 - 89.
Deninger K. 1903. Ronzotherium Reichenaui aus dem Oligocan Ton Weinheim bei Alzey. Zeitschrift der Deutschen Geologischen Gesellschaft 55: 93 - 97.
Filhol H. 1881. Etude des mammiferes fossiles de Ronzon (Haute-Loire). Annales des Sciences geologiques. 12: 1 - 270.
Gignoux M. 1928. Sur la decouverte d'un Rhinoceride (Acerotherium cf. filholi, Osborn) dans les couches petroliferes de Pechelbronn (Bas-Rhin). Bulletin du Service de la carte geologique d'Alsace et de Lorraine 1 (3): 145 - 152. https: // doi. org / 10.3406 / sgeol. 1928.1092
Heissig K. 1969. Die Rhinocerotidae (Mammalia) aus der oberoligozanen Spaltenfullung von Gaimersheim bei Ingolstadt in Bayern und ihre phylogenetische Stellung. Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse 138. Bavarian Academy of Sciences, Munich.
Heissig K. 1978. Fossilfuhrende Spaltenfullungen Suddeutschlands und die Okologie ihrer oligozanen Huftiere. Mitteilungen der Bayerischen Staatssammlung fur Palaontologie und historische Geologie 18: 237 - 288.
Jehenne Y. & Brunet M. 1992. Interet biochronologique de quelques grands mammiferes ongules de l'Eocene superieur et de l'Oligocene d'Europe. Geobios 25 (14): 201 - 206. https: // doi. org / 10.1016 / S 0016 - 6995 (06) 80329 - 3
Kafka J. 1913. Rezente und Fossile Huftiere Bohmens. (Ungulata.). Archiv fur naturwissenschaftliche Landesdurchforschung von Bohmen 14 (5): 1 - 85.
Kretzoi M. 1940. Alttertiare Perissodactylen aus Ungarn. Annales Musei Nationalis Hungarici 33: 87 - 99.
Lavocat R. 1951. Revision de la Faune des Mammiferes oligocenes d'Auvergne et du Velay. Sciences et Avenir, Paris.
Menouret B. & Guerin C. 2009. Diaceratherium massiliae nov. sp. des argiles oligocenes de Saint- Andre et Saint-Henri a Marseille et de Les Milles pres d'Aix-en-Provence (SE de la France), premier grand Rhinocerotidae brachypode europeen. Geobios 42 (3): 293 - 327. https: // doi. org / 10.1016 / j. geobios. 2008.10.009
Mermier E. 1895. Sur la decouverte d'une nouvelle espece d' Acerotherium dans la mollasse burdigalienne du Royans. Annales de la Societe linneenne de Lyon 42 (1): 163 - 190. https: // doi. org / 10.3406 / linly. 1895.4058
Osborn H. F. 1900. Phylogeny of the rhinoceroses of Europe. Bulletin of the American Museum of Natural History 13: 229 - 267.
Pictet F. J. 1853. Traite de paleontologie, ou histoire naturelle des animaux fossiles consideres dans leurs rapports zoologiques et geologiques - Seconde edition - Tome Premier. J. - B. Bailliere, libraire de l'Academie royale de Medecine, Paris. https: // doi. org / 10.5962 / bhl. title. 13903
Roman F. 1910. Sur les Rhinocerides de l'Oligocene d'Europe et leur filiation. Comptes rendus hebdomadaires des seances de l'Academie des Sciences 150: 1558 - 1560.
Roman F. 1912 a. Les Rhinocerides de l'Oligocene d'Europe. Archives du Museum d'histoire naturelle de Lyon 11: 1 - 92.
Stehlin H. G. 1909. Remarques sur les faunules de Mammiferes des couches eocenes et oligocenes du Bassin de Paris. Bulletin de la Societe geologique de France 4 (9): 488 - 520.
Uhlig U. 1996. Erstfund eines juvenilen Unterkiefers von Epiaceratherium bolcense Abel, 1910 (Rhinocerotidae, Mammalia) aus dem Unteroligozan von Monteviale (Italien). Mitteilungen der Bayerischen Staatssammlung fur Palaontologie und historische Geologie 36: 135 - 144.
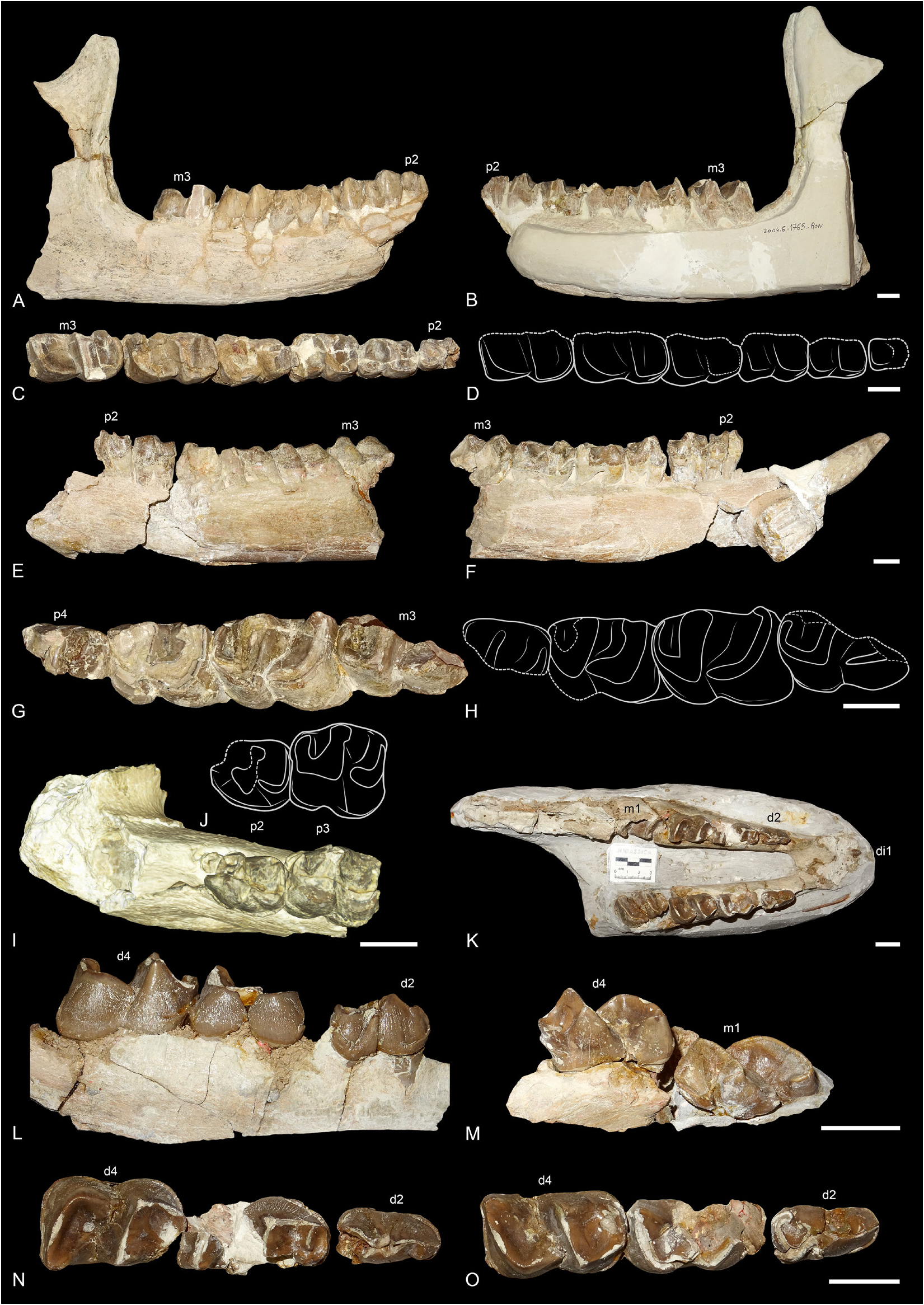
Fig. 2. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–D. Lectotype right hemimandible PUY.2004.6.1765.RON with p2–m3.A. Lateral view.B. Medial view. C. Occlusal view. D. Drawing of the occlusal view. – E–J. Broken left hemimandible PUY.2004.6.1766. RON with i2 and p2–m3. E. Lateral view. F. Medial view. G. p4–m3 in occlusal view. H. Drawing of p4–m3. I. Symphysis with p2–3 in occlusal view. J. Drawing of p2–3. – K–O. Juvenile mandible PUY.2004.7.1.RON. K. di1, d2–d4 and erupting m1 in occlusal view.L. Right d2–4 in labial view. M. Left d4–m1 in lingual view. N. Left d2–4 in occlusal view. O. Right d2–4 in occlusal view. Scale bars: 2 cm.
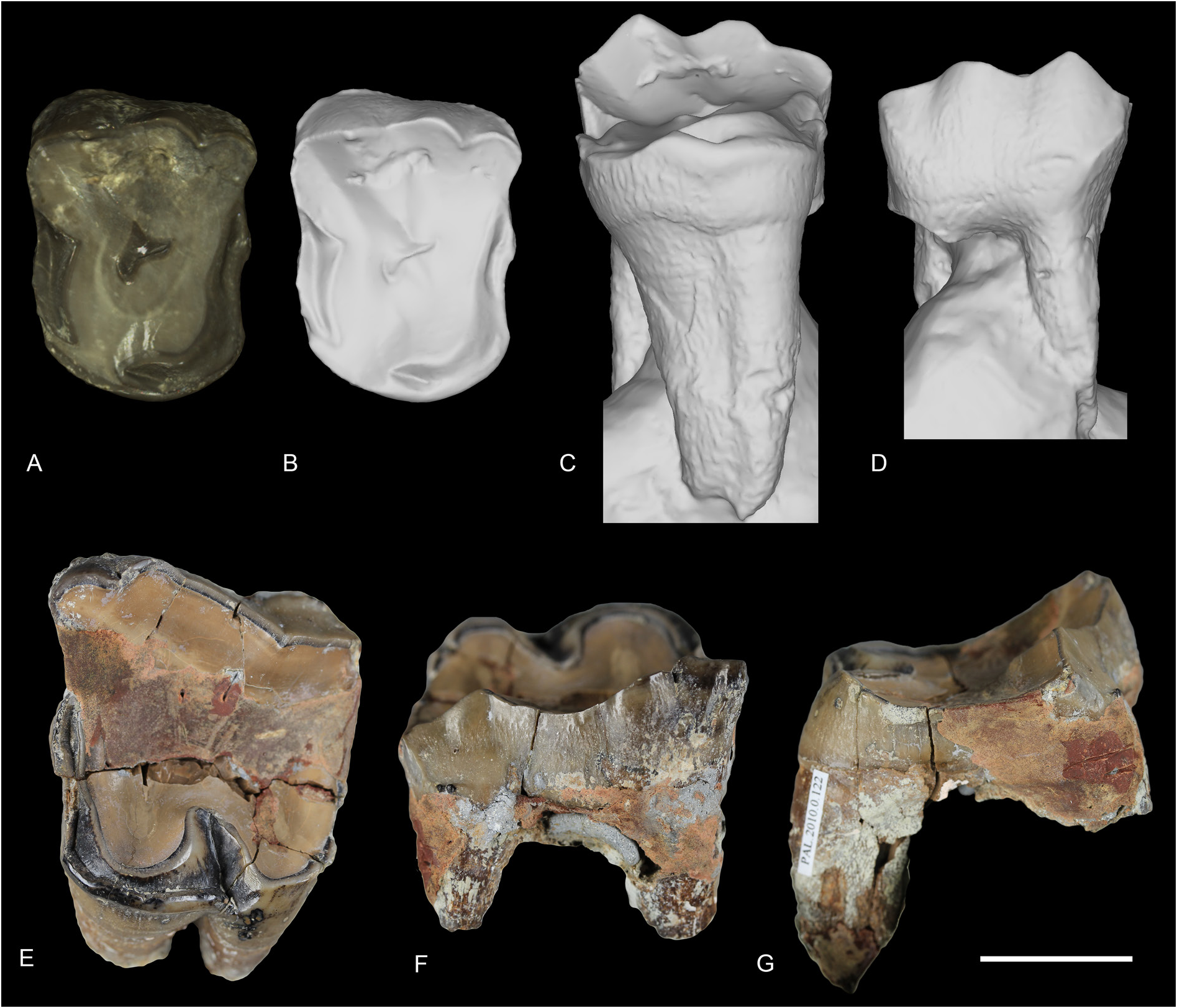
Fig. 3. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–D. 3D surface scans of P3 PUY.2004.6.1767.RON. A. With texture in occlusal view. B. Without texture in occlusal view.C. Without texture in lingual view. D. Without texture in labial view. –E–F. M1 TLM.PAL.2010.0.122. E. Occlusal view. F. Labial view. G. Lateral view. Scale bar: 2 cm.
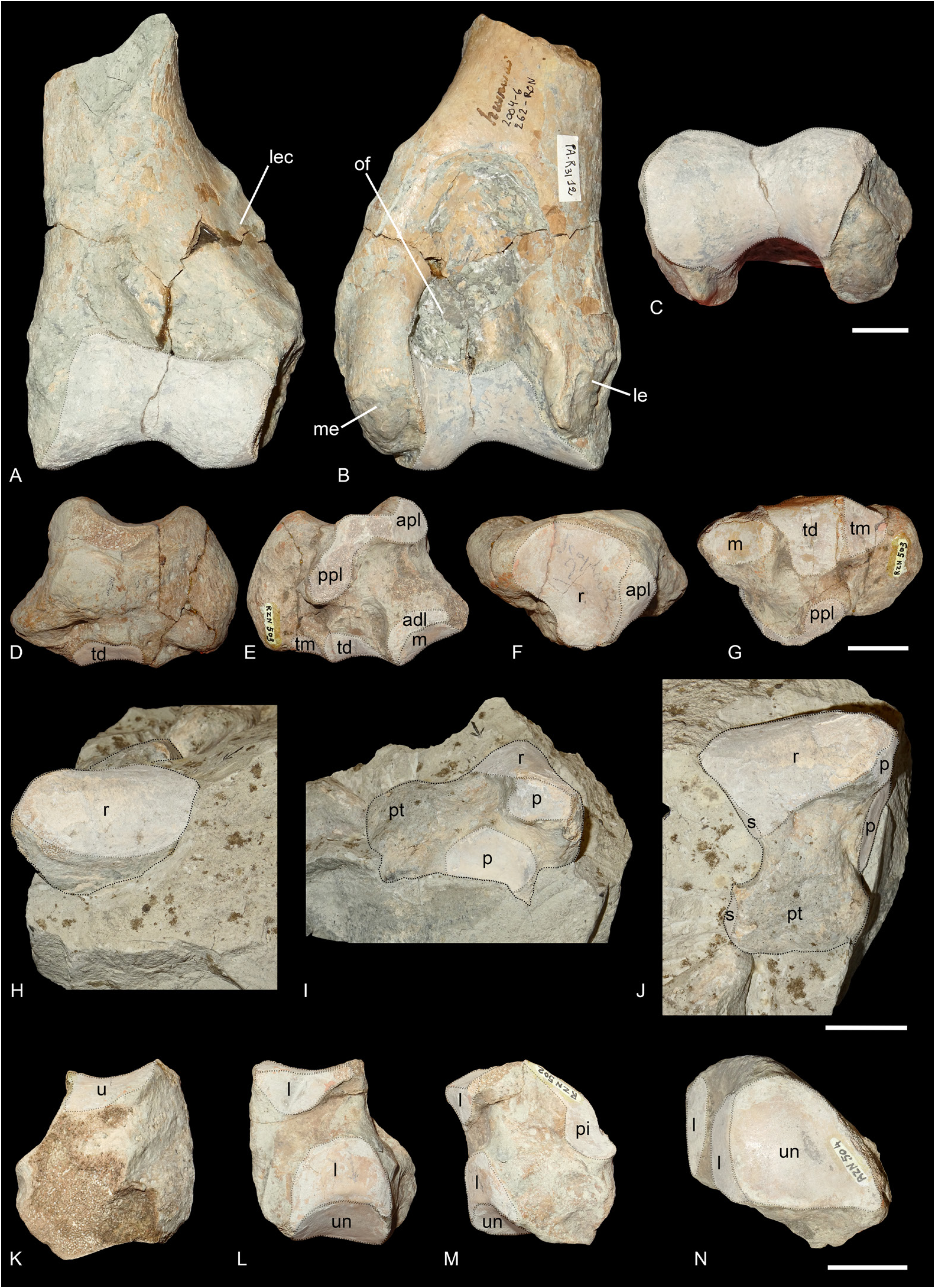
Fig. 4. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–C. Left distal humerus PUY.2004.6.262.RON. A. Anterior view. B. Posterior view. C. Distal view. – D–G. Right scaphoid MNHN.F.RZN.503. D. Medial view. E. Lateral view. F. Proximal view. G. Distal view. – H–J. Right lunate PUY.2004.6.1901.RON. H. Anterior view. I. Lateral view. J. Proximal view. – K–M. Right pyramidal MNHN.F.RZN.502. K. Lateral view. L. Medial view. M. Posterior view. – N. Left pyramidal MNHN.F.RZN.504, distal view. Abbreviations: adl = anterodistal facet for the lunate; apl = anteroproximal facet for the lunate; l = lunate; le = lateral epicondyle; lec = lateral epicondylar crest; m = magnum; me = medial epicondyle; of = olecranon fossa; p = pyramidal; pi = pisiform; ppl = postero-proximal facet for the lunate; pt = posterior tuberosity; r = radius; s = scaphoid; td = trapezoid; tm = trapezium; u = ulna; un = unciform. Articular surfaces highlighted in white. Scale bars: 2 cm.

Fig. 5. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–E. Left magnum PUY.2004.6.263.RON. A. Anterior view. B. Medial view. C. Lateral view. D. Proximal view. E. Distal view. – F–G. Left unciform PUY.2004.6.1480.RON. F. Anterior view. G. Proximal view. – H–I. Central metapodial PUY.2004.6.840.RON (possibly a MtIII) on sedimentary block, along with two probable ribs on its right and left. H. Anterior view. I. Close-up view of the proximal extremity. Abbreviations: ec = ectocuneiform; l = lunate; p = pyramidal; s = scaphoid; td = trapezoid.Articular surfaces highlighted in white. Scale bars: 2 cm.
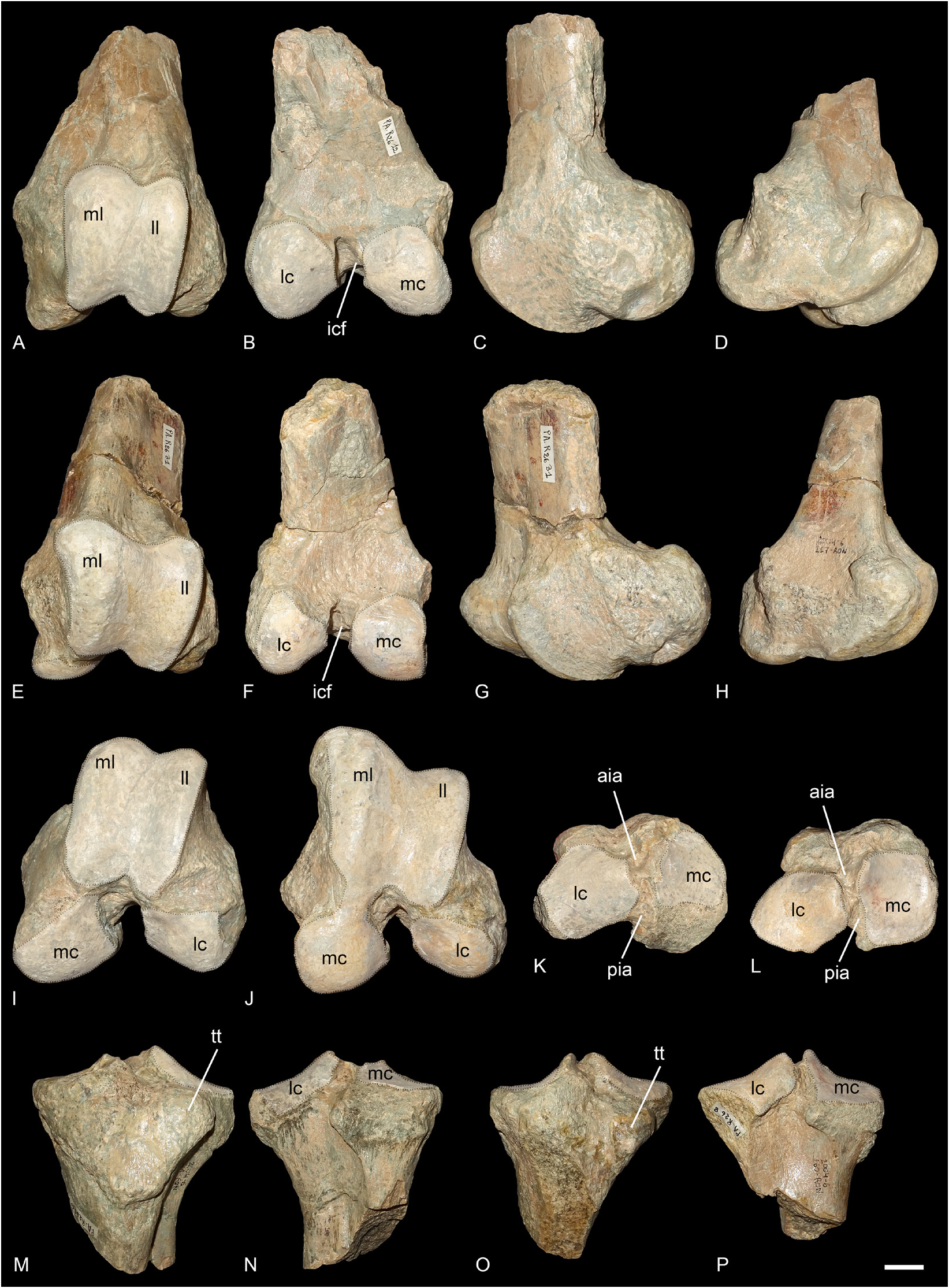
Fig. 6. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–D, I. Left distal femur PUY.2004.6.266.RON. A. Anterior view. B. Posterior view. C. Lateral view. D. Medial view. I. Distal view. – E–H, J. Left distal femur PUY.2004.6.267.RON. E. Anterior view. F. Posterior view. G. Lateral view. H. Medial view. J. Distal view. – K, M–N. Left proximal tibia PUY.2004.6.261.RON. K. Proximal view. M. Anterior view.N. Posterior view. – L, O–P. Left proximal tibia PUY.2004.6.260.RON. L. Proximal view. O. Anterior view.P. Posterior view. Abbreviations: aia = anterior intercondylar area; icf = intercondylar fossa; lc = lateral condyle; ll = lateral lip of the trochlea; mc = medial condyle; ml = medial lip of the trochlea; pia = posterior intercondylar area; tt = tibial tuberosity. Articular surfaces highlighted in white. Scale bar: 2 cm.
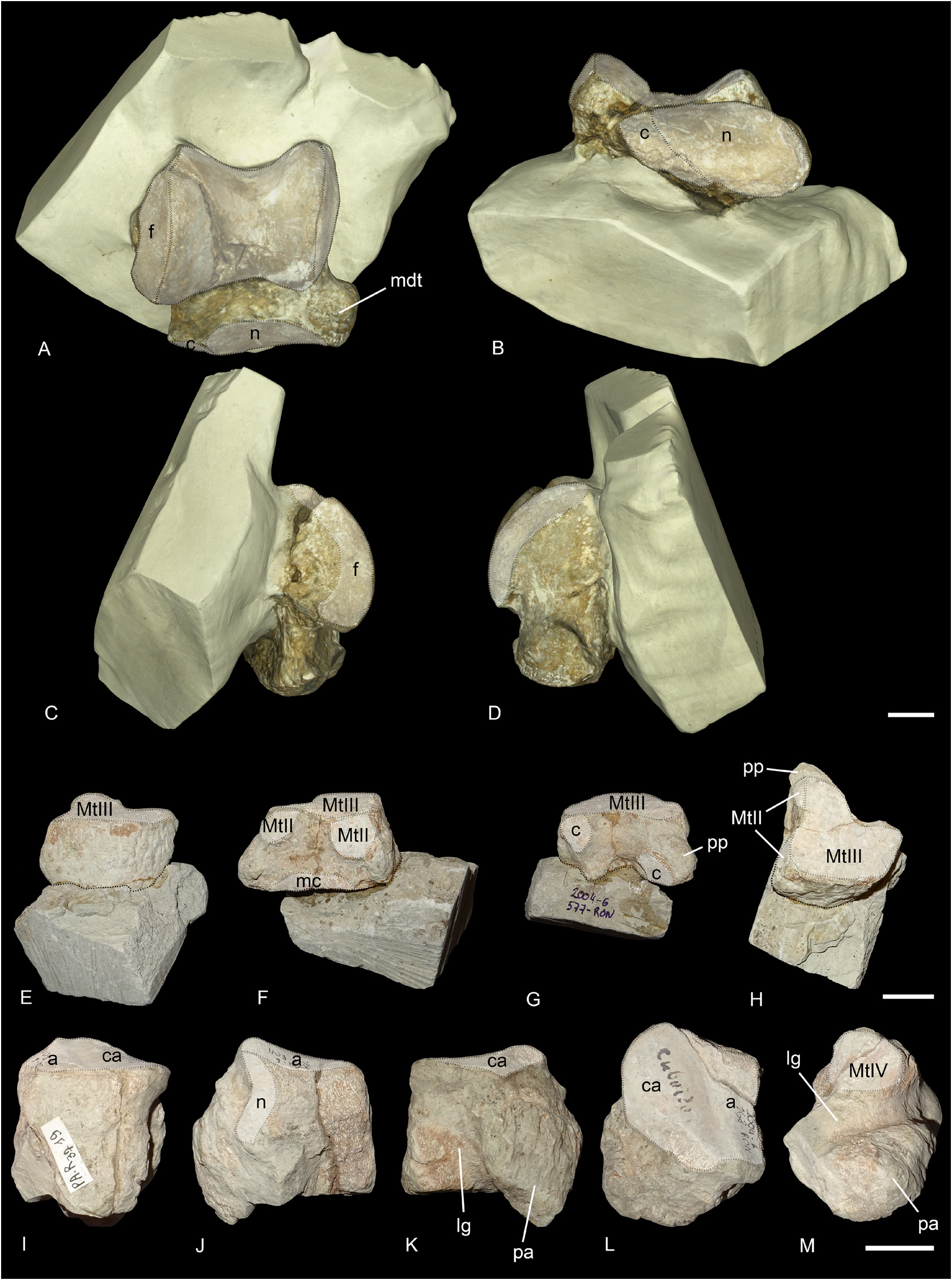
Fig. 7. Ronzotherium velaunum (Aymard in Pictet, 1853) from Ronzon (earliest Oligocene, France). – A–D. Right astragalus PUY.2004.6.1770.RON. A. Anterior view. B. Distal view. C. Lateral view. D. Medial view. – E–H. Right ectocuneiform PUY.2004.6.577.RON. E. Anterior view. F. Medial view. G. Posterior view. H. With distal border towards the top, and distal view. – I–M. Left cuboid PUY.2004.6.268.RON. I. Anterior view. J. Medial view. K. Lateral view. L. Proximal view. M. Distal view. Abbreviations: a = astragalus; c = cuboid; ca = calcaneus; f = fibula; lg = lateral groove; mc = mesocuneiform; mdt = medio-distal tubercle; n = navicular; pa = posterior apophysis; pp = posterolateral process. Articular surfaces highlighted in white. Scale bars: 2 cm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Rhinocerotoidea |
|
Family |
|
|
Genus |
Ronzotherium velaunum
| Tissier, Jérémy, Antoine, Pierre-Olivier & Becker, Damien 2021 |
Ronzotherium cf. velaunum
| Heissig K. 1978: 249 |
Ronzotherium filholi
| Lavocat R. 1951: 116 |
Acerotherium cuvieri
| Airaghi C. 1925: 26 |
Rhinoceros velaunum
| Airaghi C. 1925: 32 |
Ronzotherium velaunum
| Costeur L. & Guerin C. 2001: 77 |
| Balme C. 2000: 153 |
| Lavocat R. 1951: 115 |
| Kretzoi M. 1940: 89 |
| Kafka J. 1913: 48 |
Rhinoceros velaunus
| Roman F. 1912: 45 |
Ronzotherium cuvieri
| Filhol H. 1881: 3 |
Acerotherium velaunum
| Gignoux M. 1928: 147 |
| Roman F. 1912: 7 |
| Roman F. 1910: 1558 |
| Mermier E. 1895: 176 |
| Filhol H. 1881: 75 |
Ronzotherium cuvieri
| Aymard A. 1856: 233 |
Ronzotherium velaunum
| Menouret B. & Guerin C. 2009: 293 |
| Becker D. 2009: 495 |
| Uhlig U. 1996: 140 |
| Jehenne Y. & Brunet M. 1992: 202 |
| Brunet M. 1979: 102 |
| Brunet M. & Jehenne Y. & Ringeade M. 1977: 109 |
| Airaghi C. 1925: 25 |
| Kafka J. 1913: 5 |
| Roman F. 1912: 4 |
| Abel O. 1910: 4 |
| Stehlin H. G. 1909: 509 |
| Deninger K. 1903: 94 |
| Osborn H. F. 1900: 232 |
| Filhol H. 1881: 3 |
| Aymard A. 1856: 233 |
Rhinocéros
| Aymard A. 1854: 675 |
Acerotherium velaunum
| Pictet F. J. 1853: 296 |
Rhinoceros velaunus
| Pictet F. J. 1853: 298 |
1 (by felipe, 2021-06-14 14:00:33)
2 (by ExternalLinkService, 2021-06-14 14:07:38)
3 (by valdenar, 2021-06-15 18:27:10)
4 (by valdenar, 2021-06-15 18:32:40)
5 (by ExternalLinkService, 2021-06-15 18:51:26)
6 (by ExternalLinkService, 2021-09-19 00:56:00)
7 (by plazi, 2023-11-02 22:43:31)
8 (by ExternalLinkService, 2023-11-03 12:23:32)