
Aurumcythere tillmani Weaver & Williams
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4273.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:29AE1FBC-1E22-4A3B-B70F-97F0DB1B65D7 |
|
DOI |
https://doi.org/10.5281/zenodo.6039539 |
|
persistent identifier |
https://treatment.plazi.org/id/C53F9F72-CD77-FF9A-FF2A-FF71EF71FB31 |
|
treatment provided by |
Plazi |
|
scientific name |
Aurumcythere tillmani Weaver & Williams |
| status |
sp. nov. |
Aurumcythere tillmani Weaver & Williams , sp. nov.
Figures 2 View FIGURE 2 , 3 View FIGURE 3 & 4 View FIGURE 4
Indeterminate sp. Weaver & Williams 2016, Figs. 1 View FIGURE 1 B, 3E & F.
Type Locality. Crane Creek , at the crossing of National Forest Service Road 73, approximately 10 km north of Granite, Grant County, Oregon, USA (N 44.883317, W 118.405073). GoogleMaps
Holotype. NCSM 27006 View Materials (adult male; 339 µm length, 192 µm height); Crane Creek , Oregon, U.S.A., collected by P. G. Weaver and B. W. Williams, June 2016.
Allotype. NCSM 27000 View Materials (adult female; 385 µm length, 227 µm height); Crane Creek , Oregon, U.S.A., collected by P. G. Weaver and B. W. Williams, June 2016.
Paratypes. NCSM 27001 View Materials (adult male) and NCSM 27009 View Materials (adult female) from Crane Creek , Oregon, U.S.A. ; NCSM 27018 View Materials (adult male) and NCSM 27022 View Materials (adult male) from Olive Creek , Oregon, U.S.A. ; NCSM 27024 View Materials (adult female) and NCSM 27028 View Materials (adult male) from upper Camas Creek , Oregon, U.S.A. ; RBCM 012-00093-002 (adult male) and RBCM 012-00093-004 (adult female) from lower Camas Creek , Oregon, U.S.A. (symbiont associate of RBCM 012-00093-001 , Pacifastacus leniusculus ) ; RBCM 016-00119-003 (adult male), 016-00119-004 (adult male) and 012-00119-006 (adult female) from Desolation Creek , Oregon, U.S.A.
Diagnosis. As per genus.
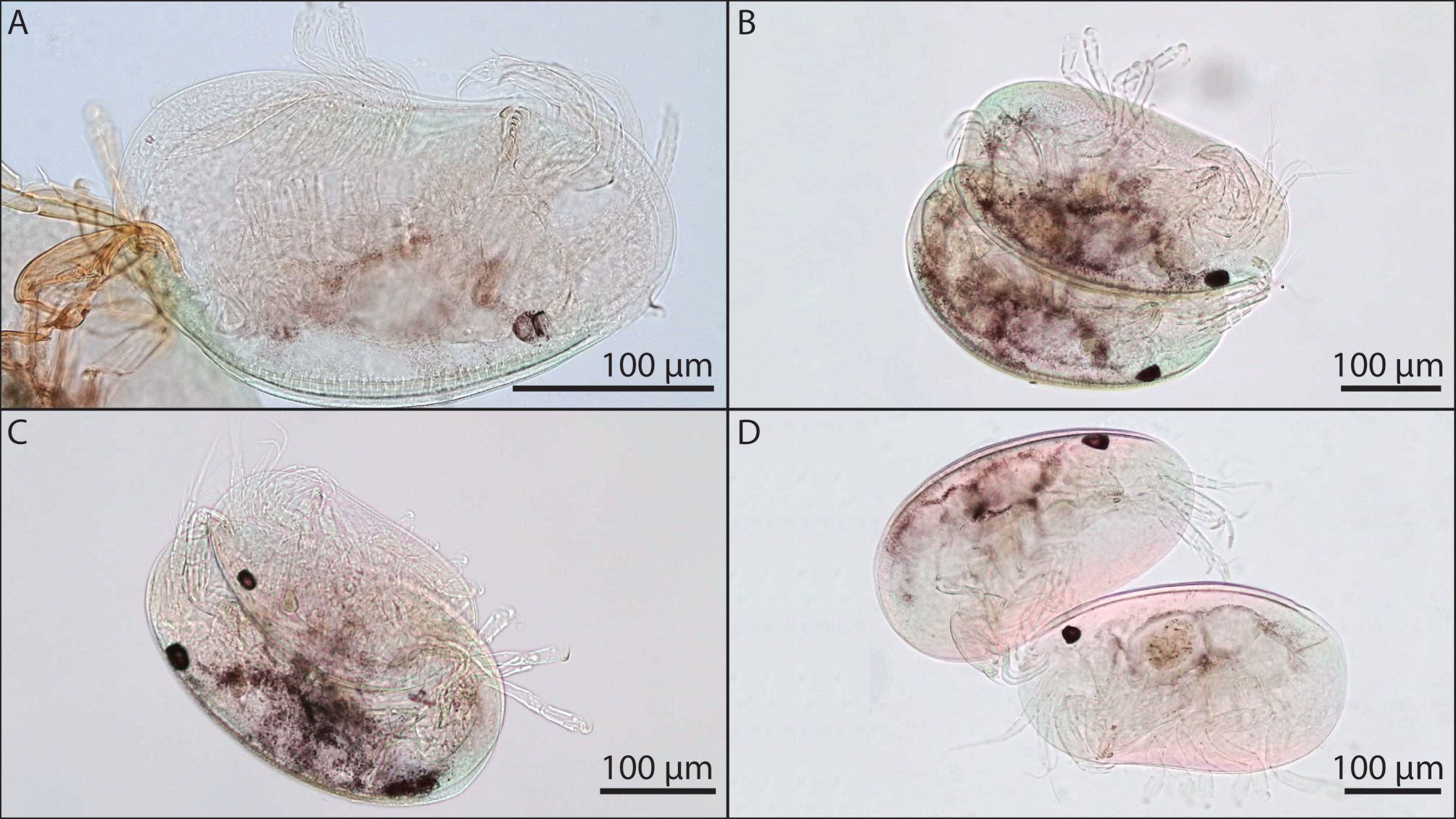
Description. ADULT MALE.—Carapace subovate to semi-elliptical ( Fig. 2 View FIGURE 2 A) with no protuberances, laterally compressed; ventral margin straight to very slightly convex, dorsal margin smoothly curved, posterior margin slightly more inflated than anterior margin; maximum height posterior to mid-length, maximum height ranging from 178–195 µm, maximum length ranging from 321–353 µm, ratio of height to length ranging from 0.52 to 0.59.
Antennule consisting of six podomeres ( Fig. 3 View FIGURE 3 A); first, i.e., most proximal, podomere without visible setae; second podomere with two setae originating from distolateral margin and extending just beyond length of third podomere; third podomere without visible setae; fourth podomere with two setae, one originating from the distal extensor margin and the other originating from the distal flexor margin; fifth podomere with two setae both extending from the distal extensor margin nearly the length of the sixth podomere; sixth podomere terminating in a cluster of five setae.
Antenna consisting of four podomeres ( Fig. 3 View FIGURE 3 B); first podomere with long, unjointed exopodite extending from distolateral margin to beyond terminal claws; second podomere with one seta extending from distal flexor margin; third podomere with one seta extending from distal flexor margin; fourth podomere without setae, bearing three terminal claws; dorsal claw with setae on distal third and notch on flexor surface proximal to setae; mesial claw short with short setae visible on distal third of flexor surface; ventral claw lacking discernable setae. Spinneret gland conspicuous, situated posterior to eyespot.
Mandible consists of palp, mandibular teeth and labial plate (sensu Hart & Hart 1974; Fig. 3 View FIGURE 3 C, D). Mandibular palp is elongate, without segmentation. Distal end of main part of palp has one small, short seta on the distal corner, and a terminal talon-shaped projection. Three long backswept setae (sensu Smith & Kamiya 2001) are discernable, extending proximally from the mid-dorsal portion of palp. There are six mandibular teeth with distal tooth larger than preceding teeth. Adjacent to mandibular teeth is a well-developed labial plate (sensu Hart & Hart, 1974) consisting of an elongate palp without segmentation, terminating in a fan-shaped segment (rake-shaped organ sensu Smith & Kamiya 2001; Fig. 3 View FIGURE 3 D).
Maxillula consists of palp, masticatory lobe and branchial plate (respiratory lobe sensu Hart & Hart 1974) ( Fig. 3 View FIGURE 3 E). Palp is elongate without segmentation, terminating in two setae. Masticatory lobe terminating in two shorter setae. Branchial plate is semi-circular with multiple setae radiating outward along its outer edge.
Fifth limb has four podomeres ( Fig. 3 View FIGURE 3 F), the first of which is broad and trapezoidal (broader proximally than distally) with two long setae extending from the distal extensor margin. Second podomere is elongate, broader distally than proximally, with one small seta on the sub-distal extensor margin. Third podomere is short and subrectangular without setae. Fourth podomere is quadrate, without setae. Distal end of fourth podomere bears a hooklike claw with five partitions.
Sixth limb has four podomeres ( Fig. 3 View FIGURE 3 G), the first of which is broad and trapezoidal (broader proximally than distally) with one long seta extending from distal extensor margin. Second podomere is elongate, slightly broader distally than proximally, with one short seta extending from sub-distal extensor margin. Third podomere is subrectangular, measures approximately half the length of the second podomere, and without setae. Fourth podomere is quadrate, without setae. Distal end of fourth podomere bears a hook-like claw with five partitions.
Seventh limb has four podomeres ( Fig. 3 View FIGURE 3 H), the first of which is elongate, broader proximally than distally, with one long seta extending from distal extensor margin. Second podomere elongate, slightly broader distally than proximally, with one short seta on sub-distal extensor margin. Third podomere sub-rectangular without setae. Fourth podomere quadrate without setae. Distal end of fourth podomere bears a hook-like claw with five partitions.
Copulatory complex of male ( Fig. 2 View FIGURE 2 C): peniferum short not extending beyond midpoint of clasping apparatus, without accessory groove and finger guard, distal end of peniferum forcipate, with posteroventral hook and spurlike prominences, both curving in an anteroventral direction, spur originating from base of hook; penis situated near distal end of peniferum; spermatic loop situated at approximately mid-height of peniferum; clasping apparatus entire moderately L-shaped, with one tooth and 5 denticles; dorsal finger, short, bulbous, inverted pear-shaped terminating in a long spine; ventral finger long, slender, approximately twice the length of dorsal finger, terminating in long spine.
Adult Female. Carapace kidney bean-shaped without protuberances ( Fig. 2 View FIGURE 2 B), laterally compressed; typically bearing an egg sac; ventral margin broadly concave medially, dorsal margin smoothly curved, posterior margin more inflated than anterior margin, maximum height slightly posterior to mid-length, maximum height ranging from 205–236 µm, maximum length ranging from 353–386 µm, ratio of height to length ranging from 0.56 to 0.62. Amiculum forcipate consisting of two roughly trianguloid structures approximately mirroring each other ( Fig. 2 View FIGURE 2 B). Antennule, mandible, maxilla and walking legs as described above for adult male. Antenna of adult female differs from that of adult male. Antenna consists of four podomeres: first podomere with long, unjointed exopodite extending from distolateral margin to beyond terminal claws; second podomere lacking setae; third podomere with one long seta extending from the distal flexor margin; fourth podomere bearing two terminal claws, and with one seta extending from mid-height on flexor margin. Antennal claws not observed to be triunguis, as is typically associated with adult females in the family Entocytheridae , but rather appear biunguis; ventral claw with setae on flexor surface; dorsal claw lacking visible setae; ventral claw without pectinate prominences at base ( Fig. 3 View FIGURE 3 I).
A-1 MALE. Carapace subovate to semi-elliptical (length 301–327 µm, height 171–185 µm), with anlagen of developing copulatory complex.
A-1 FEMALE. Carapace subovate to semi-elliptical, ventral margin mostly straight (length 321–344 µm, height 177–197 µm). Antennal claws biunguis. Bulbous projection posterior to third pair of walking legs. Nonsclerotized sub-trianguloid internal structure located in the posterodorsal portion of carapace.
Variations. Size and shape of the carapace varies throughout ontogenic and sexually dimorphic stages of development. Diagnostic characters (i.e., adult male copulatory complex, amiculum of adult female) are remarkably consistent among specimens from various localities. Although little to no variation was observed across the specimens examined during this study, perceived variation in these characters is known occur in slidemounted entocytherids depending on the angle at which a specimen is fixed in the mountant.
Variation was observed in degree of development in the copulatory complex in the anlagen of A-1 males, ranging from a largely shapeless budding structure to nearly fully-formed. The anlagen as seen in early stages of the A-1 male can be difficult to distinguish from the bulbous projection found in A-1 females; however, the A-1 female possesses a sub-trianguloid internal structure in the posterodorsal portion of carapace, a character absent in A-1 males.
Etymology. Species epithet is in honor of the late Dr. John R. Tillman, Geology Professor at Ohio Wesleyan University, Delaware, Ohio, who introduced the lead author to, and inspired in her a life-long interest in, ostracods.
Distribution. At present known only from upper reaches of the John Day River Basin in east-central Oregon, U.S.A. Aurumcythere tillmani sp. nov. was recovered from seven sites in the North Fork and Middle Fork John Day River systems ( Fig. 1 View FIGURE 1 ), including Camas Creek (N 45.058417, W 118.969917; N 45.173972, W 118.629722; N 45.047548, W 118.980912, the latter RBCM 012-00093-001), Crane Creek (type locality, N 44.883317, W 118.405073), Desolation Creek (N 44.997194, W 118.935694), Olive Creek (N 44.778972, W 118.452639), and Middle Fork John Day River (N 44.625062, W 118.582305).
Host. Pacifastacus leniusculus
Entocytherid associates. Aurumcythere tillmani sp. nov. was the only species observed from Crane Creek and Olive Creek, both small headwater streams draining into the upper North Fork John Day River. At all other sites, A. tillmani sp. nov. was found to co-occur with Uncinocythere occidentalis .
Remarks. None of the 18 adult females of A. tillmani sp. nov. examined appeared to be triunguis; all lacked the middle antennal claw associated with the final developmental stage in entocytherids. In contrast, many of the adult females of U. occidentalis examined were clearly triunguis. Rioja (1943b) noted that the middle antennal claw of Entocythere mexicana Rioja, 1943 [1943a] was transparent and inconspicuous. It is therefore possible that adult females of A. tillmani sp. nov. are in fact triunguis, but that the lack of sclerotization in A. tillmani makes it extremely difficult, if not impossible, to see the middle antennal claw of the adult female.
Aurumcythere tillmani sp. nov. appears to be unique among the nine Entocytherinae species considered native to western North America in never having sclerotized appendages, a characteristic that could be readily determined with the clearing agent and mountant used in this study. Appendages of sclerotized entocytherids appear yellowishbrown, whereas the appendages of nonsclerotized individuals—seen in most entocytherids during or immediately following molt—are clear. The lack of sclerotization appears to be characteristic of this particular species and not a function of age or molt stage, as it was consistent across all individuals identified as A. tillmani sp. nov., including copulating males. This characteristic is not likely to be a result of environment, as many of the U. occidentalis , including all copulating males, found co-occurring with A. tillmani sp. nov. had strongly sclerotized appendages. As all specimens were collected in June (either 2009 or 2016) we cannot rule out the possibility of highly synchronous molt as an explanation for the lack of sclerotization in A. tillmani sp. nov.; however, this phenomenon has never before been reported in Entocytheridae , and has not been observed in any other species examined by the authors over more extended time frames. As such, we deem this explanation unlikely, although worthy of future attention. Interestingly, the lack of sclerotization observed in A. tillmani sp. nov. may lead to higher incidences of false mating.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Entocytherinae |
|
Genus |