
Trichomycterus varii, Fernández & Bize, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4504.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:91581D02-8E8F-409C-8D60-CB953157CEA5 |
|
DOI |
https://doi.org/10.5281/zenodo.5973729 |
|
persistent identifier |
https://treatment.plazi.org/id/C27B87BC-6641-1C5D-538F-FDFABB9BF92D |
|
treatment provided by |
Plazi |
|
scientific name |
Trichomycterus varii |
| status |
sp. nov. |
Trichomycterus varii sp. nov.
Figure 1 View FIGURE 1 ; Table 1
Holotype. FACEN 0 105, 52.9 mm SL; Argentina, Jujuy Province, Santa Catalina Department, Cieneguillas River (22°07’S, 65°45’W), 3,567 m elevation, C. Fernández, 6 October 2017. GoogleMaps
Paratypes. FACEN 0 106, 9 specimens, 36.6–54.3 mm SL, collected with holotype GoogleMaps . FACEN 0 107, 2 specimens, 35.7–43.5 mm SL cs (disarticulated), collected with holotype GoogleMaps .
Diagnosis. Trichomycterus varii is diagnosed among congeners from the High Andean Plateau ( Trichomycterus alterus , T. belensis , T. boylei , T. catamarcensis , T. corduvensis , T. ramosus , T. roigi , T. yuska , T. chungaraensis , T. laucaensis , T. duellmani , T. dispar , T. therma , T. tiraquae , T. rivulatus , and T. pseudosilvinichthys ) by the caudal peduncle expanded in the area of the procurrent caudal-fin rays ( Fig. 2A View FIGURE 2 ) (vs. not expanded, Fig. 2B View FIGURE 2 ). Trichomycterus varii is further distinguished from congeners, except T. pseudosilvinichthys and T. yuska , by having the tip of pelvic fin not reaching the anus when depressed (Fig. 3A) (vs. reaching the anus Fig. 3B). Trichomycterus varii differs from T. pseudosilvinichthys by the supraorbital tendon-bone lacking a lateral process (vs. lateral process present), 15–19 interopercular odontodes (vs. 32–42), 16 pairs of ribs (vs. 17–18), 25 caudal vertebrae (vs. 26–29), 36 total vertebrae (vs. 37–40), the supraorbital sensory pore s3 present (vs. absent) and from T. yuska by having the first pectoral-fin ray ending at the margin of the fin (vs. prolonged as a short distal filament), the supraorbital tendon-bone lacking a lateral process (vs. lateral process present), 15–19 interopercular odontodes (vs. 25–29), 16 pairs of ribs (vs. 18–19), 25 caudal vertebrae (vs. 27–30), 36 total vertebrae (vs. 38–39), the supraorbital sensory pore s3 present (vs. absent), the infraorbital sensory pores i1 and i3 present (vs. absent). Trichomycterus varii is further distinguished from T. alterus by the head rectangular in dorsal view (vs. triangular), the maxillary barbel narrowed basally (vs. widened), the interopercular odontodes not embedded in fleshy tissue (vs. embedded in thick integument), the first pectoral-fin ray ending at the margin of the fin (vs. prolonged as a short distal filament), the origin of the dorsal fin pigmented (vs. unpigmented), the caudal fin margin truncated (vs. emarginated), the premaxilla rectangular in ventral view (vs. oval), the supraorbital tendon-bone without lateral process (vs. lateral process present), 13 principal caudal-fin rays (vs. 12), 11 precaudal vertebrae (vs. 3–5), 25 caudal vertebrae (vs. 31–34), 16 pairs of ribs (vs. 11–14); from T. belensis by the head rectangular in dorsal view (vs. triangular), the interopercular odontodes not embedded in fleshy tissue (vs. embedded in thick integument), the origin of the dorsal fin pigmented (vs. unpigmented), the absence of a vertical dark bar across the caudal-fin base (vs. vertical bar present), the supraorbital tendon-bone without lateral process (vs. lateral process present), the premaxilla rectangular in ventral view (vs. oval), 25 caudal vertebrae (vs. 29–32);from T. boylei by the caudal fin margin truncated (vs. emarginated), the skin of trunk with minute thread-like papillae (vs. with large papillae), the supraorbital tendon-bone without lateral process (vs. lateral process present), the premaxilla rectangular in ventral view (vs. oval); from T. catamarcensis by the presence of the pelvic fin and girdle (vs. absent), 7 pectoral fin rays (vs. 8); 15–19 interopercular odontodes (vs. 23–30), the supraorbital canal with sensory pore 3 (vs. absent), 36 vertebrae (vs. 37–39), 16 pairs of ribs (vs. 18–20); from T. corduvensis by the supraorbital canal with sensory pore 3 (vs. absent), the skin of trunk with minute thread-like papillae (vs. with large papillae), the laterosensory canal of trunk with three sensory pores (vs. 5–6), the premaxillary teeth conical, not expanded distally (vs. spatulate and distally expanded); from T. ramosus by having simple unbranched barbels (vs. barbels distally branched one or more times), the supraorbital tendon-bone without lateral process (vs. lateral process present), the premaxilla rectangular (vs. oval), 36 vertebrae (vs. 38–39); from T. laucaensis by the head approximately rectangular in dorsal view (vs. triangular), 15–19 interopercular odontodes (vs. 34–60), 7 pectoral fin rays (vs. 8–9); 36 vertebrae (vs. 38–42); from T. chungaraensis by the tip of nasal barbel not reaching the opercular (vs. reaching the opercular), 15–19 interopercular odontodes (vs. 45–55), 7–14 opercular odontodes (vs. 14–20), 7 pectoral fin rays (vs. 8–9); 36 vertebrae (vs. 37–40); from T. duellmani by the laterosensory canal of trunk with three sensory pores not enclosed by ossicles (vs. 4 pores with 5–6 ossicles), the marmorated pigmentation on lateral body (vs. two longitudinal dark lines), 7 pectoral fin rays (vs. 8–9), 15–19 interopercular odontodes (vs. 32–40), 16 pairs of ribs (vs. 12–13); from T. therma by the thickened transverse skin on the ventral surface of the head absence (vs. presence), the laterosensory canal of trunk with three sensory pores (vs. 4–6), 7 pectoral fin rays (vs. 9), 15–19 interopercular odontodes (vs. 20–27), 36 total vertebrae (vs. 32), 16 pairs of ribs (vs. 12); T. tiraquae by the supraorbital canal with sensory pore 3 (vs. absent), the skin of trunk with minute thread-like papillae (vs. with large papillae), the laterosensory canal of trunk with three sensory pores (vs. 5–6); T. rivulatus by 7 pectoral fin rays (vs. 8–9), 15–19 interopercular odontodes (vs. 31–40), 36 total vertebrae (vs. 37–39); T. dispar by the first pectoral-fin ray terminating at fin margin without forming distal filament (vs. prolonged as a short distal filament). T. varii further differs from the sympatric T. roigi by 7 pectoral fin rays (vs. 9), 15–19 interopercular odontodes (vs. 38–40).
Description. Morphometric data for holotype and paratypes given in Table 1. Body elongate, cylindrical overall in trunk region and gradually and progressively becoming more compressed towards caudal fin. Dorsal and ventral profiles of trunk slightly straight. Caudal peduncle expanding posteriorly toward caudal fin. Papillae-like structures present on trunk of body. Urogenital opening closer to anal-fin origin than to pelvic-fin insertion and pelvic fin when depressed not covering anus.
TABLE 1 . Morphometric data for holotype and 9 paratypes of Trichomycterus varii , new species. Standard length and head length are expressed in millimeters; measurements 1 to14 are percentages of standard length; 15 to 23 are percentages of head length .
Head shape approximately rectangular in dorsal view. Head dorsoventrally flattened with eye located on dorsal surface and not visible in lateral view. Anterior profile of head semicircular. Adductor musculature of cheek well developed. Skin covering eye thin, transparent, and separate from surface of eyeball.
Anterior nostril slightly smaller than posterior nostril and bordered medially by fleshy flap and laterally by base of nasal barbel. Flap and base of barbel continuous and forming short tube. Anterior margin of posterior nostril bordered by flap of thin skin.
Mouth subterminal, with rictus directed posteriorly. Upper lip fleshy with numerous papillae. Lower lip with prominent fleshy lobes along lateral margin; lobes situated internal to base of rictal barbels. Lower lip fleshy, with papillae-like structures covering anterior and to lesser degree anteroventral surfaces.
Barbels short and tapering distally, but not thread-like or with distal branching. Tip of maxillary barbel falling short of vertical through anterior limit of patch of opercular odontodes. Submaxillary barbel shorter than maxillary barbel and falling short of vertical through anterior limit of opercular patch of odontodes. Nasal barbel slender, reaching posteriorly distinctly beyond posterior border of eye. Origin of nasal barbel situated on posterolateral portion of skin flap running along margin of anterior naris.
Branchiostegal membrane narrowly attached to isthmus anteriorly at midline, branchial opening wide. Branchiostegal rays six or seven in two cs specimens. Interopercular patch of odontodes elongate, with maximum two irregular rows of odontodes. Fleshy interopercular less developed than in congeners, with 12 to 15 odontodes visible in alcohol specimens and 15 to 19 odontodes present in two cs specimens. Opercular odontodes not deeply embedded in fleshy tissue on opercle. Opercular patch of odontodes somewhat rounded, with five to 12 straight odontodes obvious in alcohol specimens and seven to 14 odontodes present in two cs specimens.
Mesethmoid T -shaped, elongate; anterior margin straight; shaft smaller than lateral cornua, its posterior process extending between lateral ethmoids, vomer, and anterior portion of frontals. Anterior cranial fontanel reduced to
Supraorbital canal with pores s1, s2, s3, and s6. Infraorbital canal incomplete, with segment between pores i3 and i10 absent. Posterior portion of infraorbital canal situated posterior to rim of orbit and arising from temporal canal running within sphenotic. Posterior portion of infraorbital canal with infraorbital pores i10 and i11. Postotic canal with two pores, with pterotic branch present at junction of pterotic and posttemporo-supracleithrum. Laterosensory canal of trunk reduced, with three pores
Dorsal-fin rays 12 to 13, with six to seven branched and six unbranched rays in two cs specimens; one unbranched and six to seven branched rays externally visible with remaining rays hidden under thick integument. Distal margin of dorsal fin rounded. Dorsal-fin origin located distinctly anterior to vertical through anterior limit of vent, midway between pelvic and anal-fin origins. Anal-fin rays nine, with five branched and four unbranched rays in two cs specimens; externally visible rays one unbranched and five branched with remaining rays hidden under thick integument. Anal fin relatively elongated, equal in size to, or slightly smaller than dorsal fin, with distal margin slightly rounded. Anal-fin origin located approximately at vertical through middle or posterior portion of base of dorsal-fin base. Pectoral-fin rays seven, with lateral-most ray unbranched. Second and third pectoral-fin rays longest. Distal margin of pectoral fin almost rounded. First pectoral-fin ray terminating at fin margin without forming distal filament. Pelvic-fin rays five, with four branched and one unbranched rays, plus small splint. Second and third pelvic-fin rays longest. Tip of pelvic fin when depressed not reaching posteriorly anal opening. Pelvic fin with thick adipose layer in small specimens. Distal margin of caudal fin nearly truncated. Principal caudal-fin rays 6+7. Caudal peduncle markedly expanded in area of procurrent rays but never deeper than caudal-fin base. Dorsal procurrent caudal-fin rays 10 to 11 in two cs specimens. Ventral procurrent caudal-fin rays 11 in two cs specimens. Total vertebrae 36, with 11 precaudal and 25 caudal vertebrae. Ribs on each side of body 16. No external obvious sexual dimorphism present.
Pigmentation in alcohol. Trichomycterus varii demonstrates considerable range in the intensity of dark pigmentation on the head and body. The holotype is relatively marmorated, while several paratypes are lightly pigmented on the head and trunk. Supraorbital and infraorbital canal are unpigmented in some paratypes. All specimens slightly pigmented on ventral surface of head and trunk. Dorsal, anal, pectoral and caudal fins irregularly marmorated and pelvic fin lightly pigmented. All barbels darkly pigmented at least to some degree. Opercular and interopercular odontodes unpigmented.
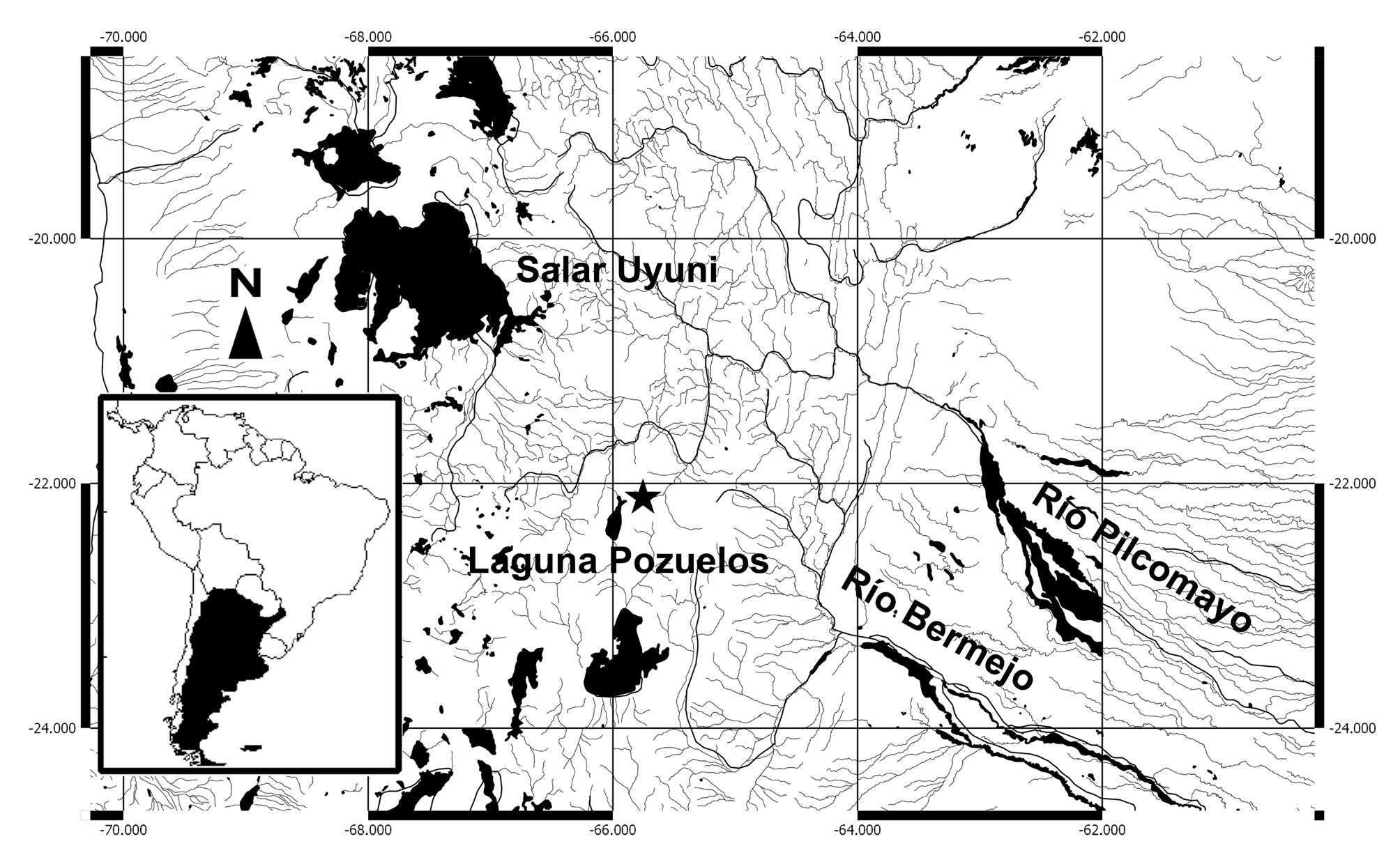
Ecology and endemicity. The type locality of Trichomycterus varii is a shallow (0.10-0.40 m) stream over different types of substrates (rocks, gravel, sand, and decomposing organic material) ( Fig. 4 View FIGURE 4 ). Stomachs of two cs specimens of T. varii contained dipteran larvae ( Chironomidae and Ceratopogonidae ) and filamentous algae. The only other species of fish collected at that site is T. roigi . Sampling efforts in adjoining drainage systems failed to yield any specimens of T. varii , which consequently appears to be endemic to the Río Cieneguillas. Members of Trichomycterus and Silvinichthys seem well adapted to the high elevation, temporary endorrheic or poorly drained basins, and occasionally permanent watercourses fed by snow melt of higher elevations; all of which are typical habitats of the High Andean Plateau.
Distribution. Trichomycterus varii is only known from the type locality ( Fig. 5 View FIGURE 5 ) that lies within a region with few drainage systems and a depauperate ichthyofauna. Trichomycterus varii is the sixth fish species apparently endemic to the High Andean Plateau from Argentina ( Fernández & Vari 2012; Fernández & Andreoli Bize 2017: Fig. 2 View FIGURE 2 ) and the seventeenth species known to occur at higher elevations (at or above 3,000 m); the other species are T. chungaraensis Arratia 1983 , T. laucaensis Arratia 1983 from Chile, T. duellmani Arratia & Menu-Marque 1984 , T. therma Fernández & Miranda 2007 , T. tiraquae ( Fowler 1940) , T. pseudosilvinichthys Fernández & Vari 2004 from Bolivia, T. dispar ( Tschudi 1846) from Perú, T. rivulatus Valenciennes, 1846 from Bolivia and Perú, T. alterus ( Marini, Nichols & La Monte 1933) , T. boylei ( Nichols 1956) , T. corduvensis Weyenberg 1877 , T. catamarcensis Fernández & Vari 2000 , T. ramosus Fernández 2000 , T. belensis Fernández & Vari 2002 , T. roigi Arratia & Menu- Marque 1984, and T. yuska Fernández & Schaefer 2003 from Argentina. Unfortunately, the knowledge of the biology and ecology of these species is still incomplete.
Etymology. Trichomycterus varii is named after Richard Peter Vari for his outstanding contribution to the knowledge of the South American freshwater fishes and especially from Andes mountains.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |