
Iscini, Kluge, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3760.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:A180D4A6-78B7-4FFB-A291-728B2FAF3495 |
|
DOI |
https://doi.org/10.5281/zenodo.5040933 |
|
persistent identifier |
https://treatment.plazi.org/id/C0488791-BA7A-AC35-F5EA-FEE972C308E3 |
|
treatment provided by |
Felipe |
|
scientific name |
Iscini |
| status |
trib. nov. |
Iscini , tribus novum
Type genus: Isca Gillies 1951 .
Typified name in basic format: Isca /fg (incl. Notophlebia ).
Belongs to Atalophlebolinguata Kluge 2009 (or Atalophlebiinae in the narrowest sense), and has all apomorphies of this taxon ( Kluge 2009, 2012). Other characters of Iscini are listed below; among them, characters (1), (2), (5), (7) and (10) are autapomorphies which are not found in other taxa and testify about holophyly of Iscini .
In larva:
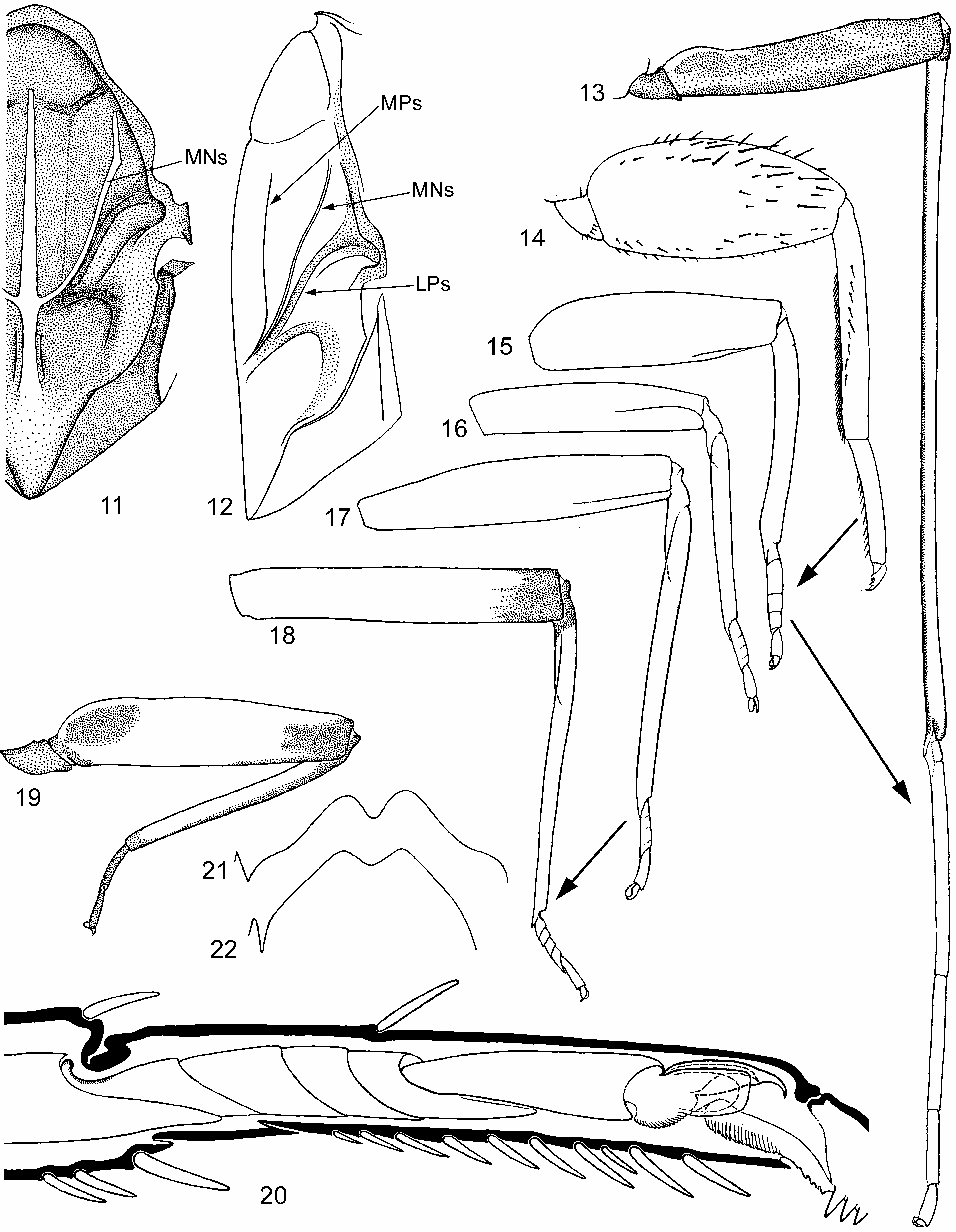
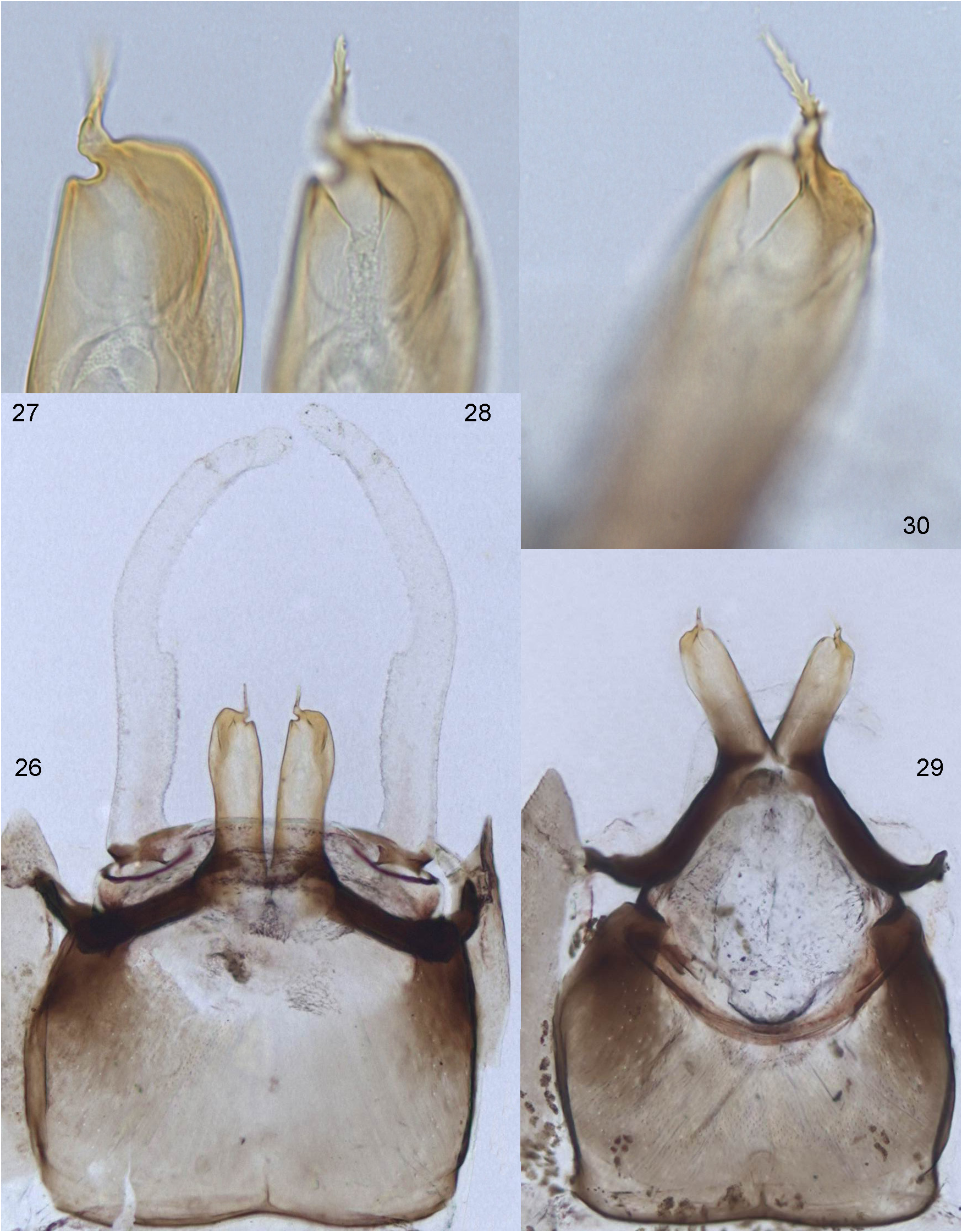
(1) On maxilla apical-ventral row of pectinate setae (which is characteristic for Leptophlebiidae ) is unusually short and consists of 3 setae; these setae are retained in Isca and in Notophlebia ganeshi sp.n. ( Fig. 6 View FIGURES 4–10 ), but are lost in Notophlebia jobi due to its modified dilatognathan mouth apparatus ( Sivaramakrishnan & Peters 1984: Fig. 15 View FIGURES 11–20 ).
Among other Leptophlebiidae , reduction of these setae occurs only in some representatives with dilatognathan mouth apparatus: three vestigial setae are present in Dilatognathus Kluge 2012 ; these setae are completely lost in Hermanellognatha ( Kluge 2012). In other known leptophlebiid taxa (including dilatognathan taxa Hagenulus Eaton 1882 s.str. and Ulmeritus Traver 1956 s.str.) there is a row of many well-developed pectinate setae.
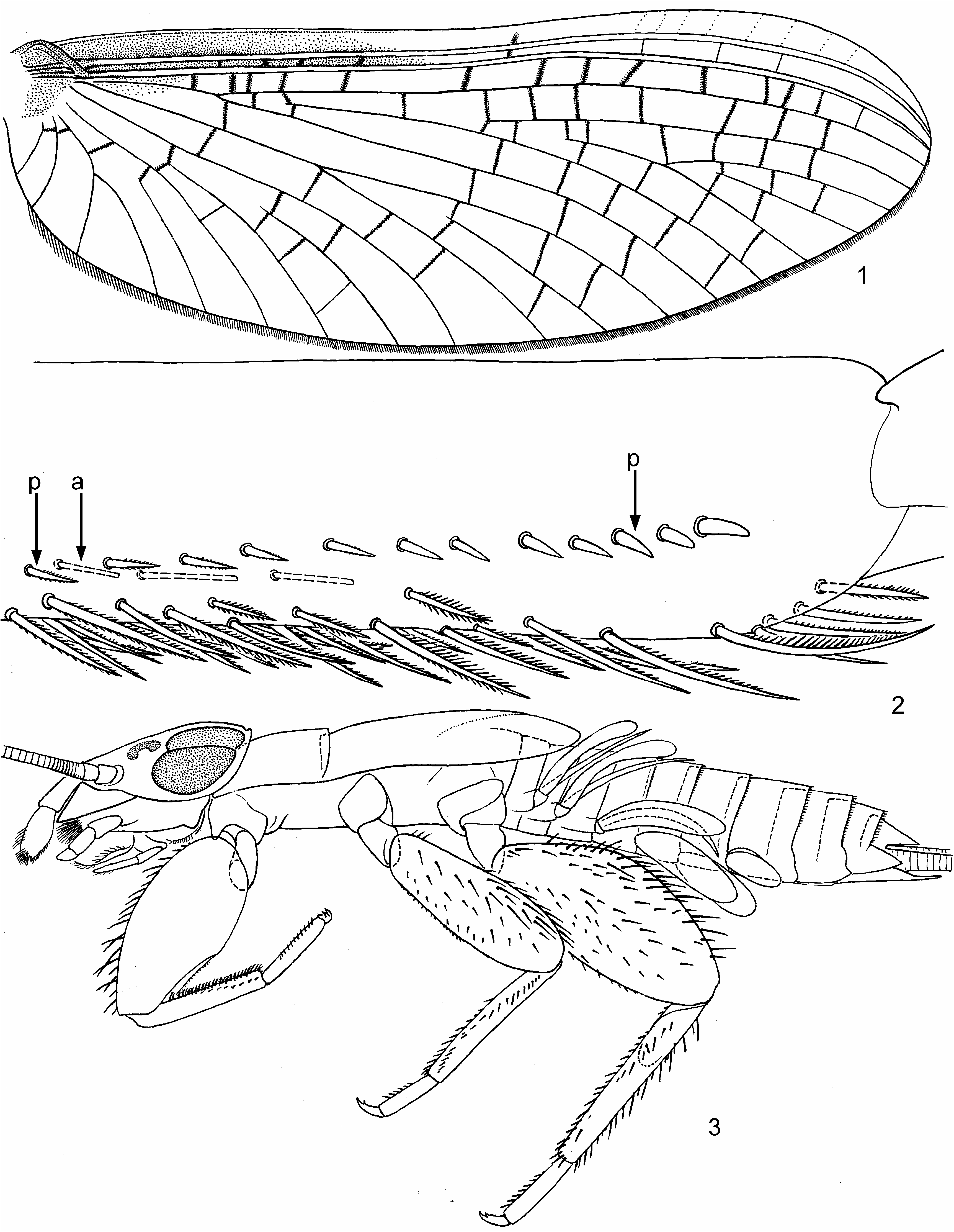
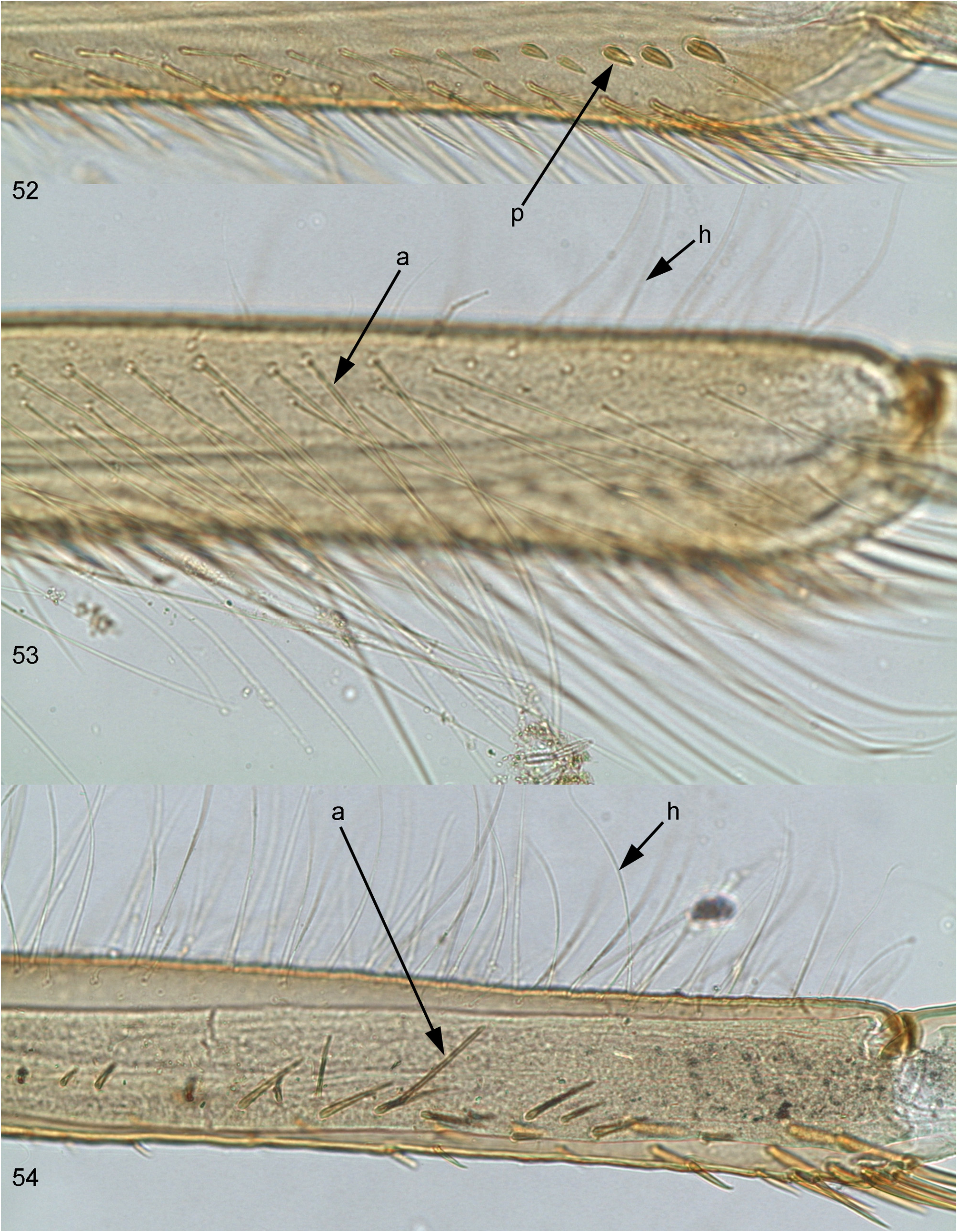
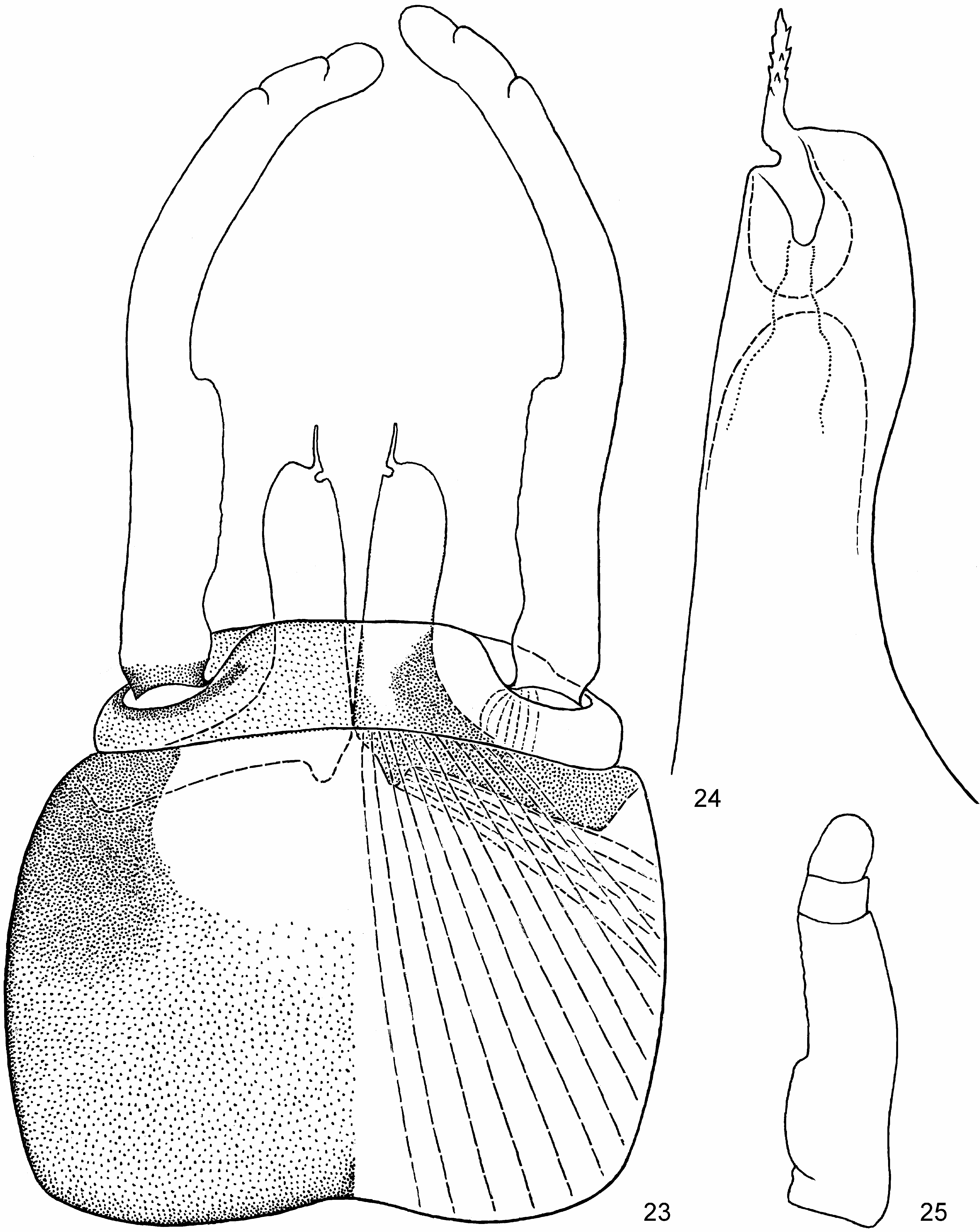
(2) On larval fore tibia field of stout pointed bipectinate setae on inner side (which is characteristic for Leptophlebiidae ) gradually turns to a longitudinal row of stout non-pectinate setae, located in distal part of ventral (posterior) side of tibia; most distal seta of this row is the most stout ( Figs 2 View FIGURES 1–3 , 52p View FIGURES 52–54 ).
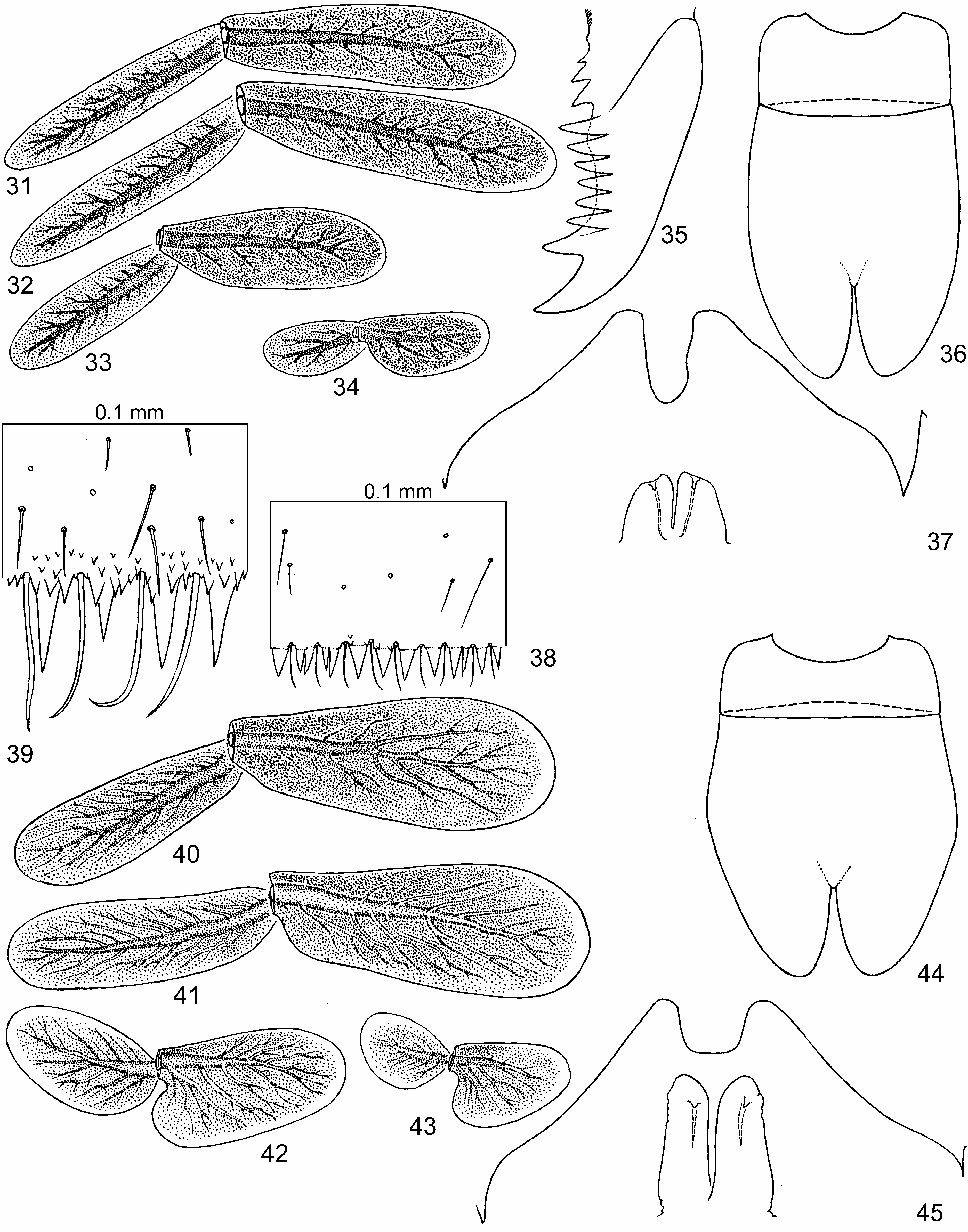
(3) In larva, patella-tibial suture (initially absent on fore legs) is lost on middle leg, being retained on hind leg only ( Fig. 3 View FIGURES 1–3 ); unlike larva, in subimago and imago patella-tibial suture is retained both on middle and hind legs ( Figs 16–18 View FIGURES 11–20 ). The same in some other taxa (e.g., Megaglena Peters & Edmunds 1970 ).
(4) Larval claw has a row of subequal denticles and one larger distal denticle located posteriad of this row ( Fig. 35 View FIGURES 31–45 ). The same in some other taxa.
(5) While insertions of all tergalii retaine their position on posterior-lateral angles of abdominal terga, terga become gradually wider and sterna become gradually narrower from abdominal segment I to abdominal segment VII, so that more posterior tergalii are inserted gradually more ventrally, than more anterior ones. In Notophlebia insertions of tergalii I–VI are gradually shifted from dorsal to lateral sides of abdomen; tergalii VII are lost ( Fig. 3 View FIGURES 1–3 ). In Isca tergalii I are lost, and insertions of tergalii II–VII are gradually shifted from lateral to ventral sides of abdomen. Both in Notophlebia and Isca , abdomen is cylindric, with terga strongly arched and rigid. Anterior abdominal segments are somewhat shorter than posterior ones, so that tergalii insertions are somewhat brought together.
In adult:
(6) Adults are shortly-molting: both subimaginal and imaginal cuticles are developed before larval/ subimaginal molt ( Fig. 20 View FIGURES 11–20 ), and subimago molts to imago shortly after emergence. The same in some other mayfly taxa.
(7) Pigmented areas, which initially for Leptophlebiidae are present on cuticle of subimaginal mesonotum ( Kluge 2004), are present on cuticle of imaginal mesonotum (in contrast to other Leptophlebiidae , whose cuticle of imaginal mesonotum is evenly pigmented). The pair of pigmented areas bordered by mesonotal suture are enlarged and occupy most part of medioscutum and submedioscutum, so that only three narrow colorless stripes separate them one from another and from other pigmented areas of mesonotum ( Fig. 11 View FIGURES 11–20 ). In Isca such pigmentation occurs both on imaginal and subimaginal cuticle. In Notophlebia ganeshi sp.n. subimaginal cuticle is entirely colorless ( Fig. 12 View FIGURES 11–20 ).
(8) Hind wings are completely lost; larval metanotum lacks vestiges of protoptera ( Fig. 3 View FIGURES 1–3 ). The same in some other taxa.
(9) Subimaginal legs in some degree retain size and proportions of larval legs: subimaginal fore femur is widened proximally, and hind femur widened in middle ( Figs 15–17 View FIGURES 11–20 ). Fore tibia and tarsus of male subimago are as short as in larva, and tarsal segments II–IV are swollen ( Fig. 15 View FIGURES 11–20 ); after molt to imago, tibia and tarsus become several times longer ( Fig. 13 View FIGURES 11–20 ). In most other leptophlebiids changes of male fore leg at molt from subimago to imago is not so strong.
(10) On middle and hind leg, 1 st tarsal segment (initially for Ephemeroptera fused with tibia, and initially for Furcatergaliae strongly shortened) is secondarily separated from tibia and elongated ( Fig. 18 View FIGURES 11–20 ). On inner side of leg, apex of tibia is stretched distally and retains fusion with 1 st tarsal segment; on outer side of leg, 1 st tarsal segment is well separated from tibia and proximally has a concavity, which corresponds to condylus in tibio-tarsal articulation of the larval cuticle, under which subimaginal and imaginal leg had been developed ( Fig. 20 View FIGURES 11–20 ). Thus, number of distinct tarsal segments can be 5 (instead of 4 in most other Leptophlebiidae ). 1 st, 2 nd, 3 rd and/or 4 th tarsal segments are short and can be more or less fused one with another; sometimes some of them are fused completely. Thus, total number of distinguishable tarsal segments can be either 5 (e.g., in Notophlebia ganeshi sp.n. and Isca purpurea Gillies 1951 ) or diminished to 4 or 3 (e.g., in Isca serendiba Peters & Edmunds 1970 ). In female, structure of fore tarsus ( Fig. 19 View FIGURES 11–20 ) is similar to that of middle and hind tarsus. About fore tarsus of male see above (8).
(11) Imaginal and subimaginal claws of all legs are ephemeropteroid. Plesiomorphy ( Kluge 2004).
(12) Penis consists of a pair of L-shaped sclerotized lobes movably connected basally ( Fig. 23 View FIGURES 23–25 , 26, 29 View FIGURES 26–30 ). The same in some other taxa.
(13) Caudalii of subimago (which in mayflies are usually developed from proximal portions of larval caudalii) are developed from very small proximal portions of larval caudalii, so that are several times shorter than larval caudalii; caudalii of male imago have strongly elongated segments, so that are several times longer than subimaginal caudalii; caudalii of female imago in Notophlebia ganeshi sp.n. are subequal to subimaginal ones (in Isca female subimago does not molt to imago). In other leptophlebiids degeneration and growth of caudalii during metamorphosis are not so great.
Distribution and composition. Oriental Region. Includes Isca Gillies 1951 and Notophlebia Peters & Edmunds 1970 .
Discussion. Peters & Edmunds (1970) ascribed the character (1) to their lineage IIB5, which included Isca , Notophlebia and Nathanella . This character was formulated as "A row of thick spines absent on ventral surface of maxillae near inner anterolateral margin (Figs 193–195, 350)". Actually this row of stout pectinate spine-like setae (not spines) is not absent, but reduced to 3 setae ( Fig. 6 View FIGURES 4–10 ; Peters & Edmunds 1970: Fig. 350) and can be overlooked ( Peters & Edmunds 1970: Figs 194, 195); only in Notophlebia jobi it is really absent ( Peters & Edmunds 1970: Fig. 193). At that time larvae of Nathanella had not been discovered, and larva of Notophlebia jobi was erroneously ascribed to Nathanella . Actually, Nathanella has this row of pectinate spine-like setae well developed ( Sivaramakrishnan et al. 1996: Fig. 16 View FIGURES 11–20 ). Within the line IIB5, Peters & Edmunds (1970) accepted daughter line IIB5a, which united Notophlebia with Nathanella and differed from Isca only by plesiomorphy—evenly arranged cross veins. Uniting of Notophlebia with Nathanella , being based solely on symplesiomorphy, is not grounded.
Here the formal tribus rank is ascribed to this taxon. However, number of generally accepted formal familygroup ranks is not enough for phylogenetic classification of Leptophlebiidae . Between the taxon Leptophlebiidae , whose family rank is generally accepted, and the tribe Iscini , there are subsequently subordinated holophyletic taxa with non-typified circumscriptional names Atalophleboadentata, Atalophlebopectinata, Atalophleboculata, Atalophlebomaxillata and Atalophlebolinguata ( Kluge 2009); at the same time, the tribe Iscini includes taxa Isca and Notophlebia , whose generic ranks are generally accepted. Recently, position of Iscini within Atalophlebolinguata remains to be unclear; possibly, after its clarification more hierarchical levels will be established. At the present moment, when the taxon named here Iscini , includes only two genera — Isca and Notophlebia , the rank-based name Iscini can be used for it; if number of subsequently subordinated taxa with typified name Isca /fg will be increased, this rank-based name can become inconvenient and should be substituted by a hierarchical name.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |