
Pseudachorutes concinnus Babenko & Nakamori, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4938.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:6FEECE37-B2D3-4AE3-9878-CF212420AF9C |
|
DOI |
https://doi.org/10.5281/zenodo.4605156 |
|
persistent identifier |
https://treatment.plazi.org/id/BF05878D-232F-FFE0-FF49-899BFD6CCD8A |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudachorutes concinnus Babenko & Nakamori |
| status |
sp. nov. |
Pseudachorutes concinnus Babenko & Nakamori sp. nov.
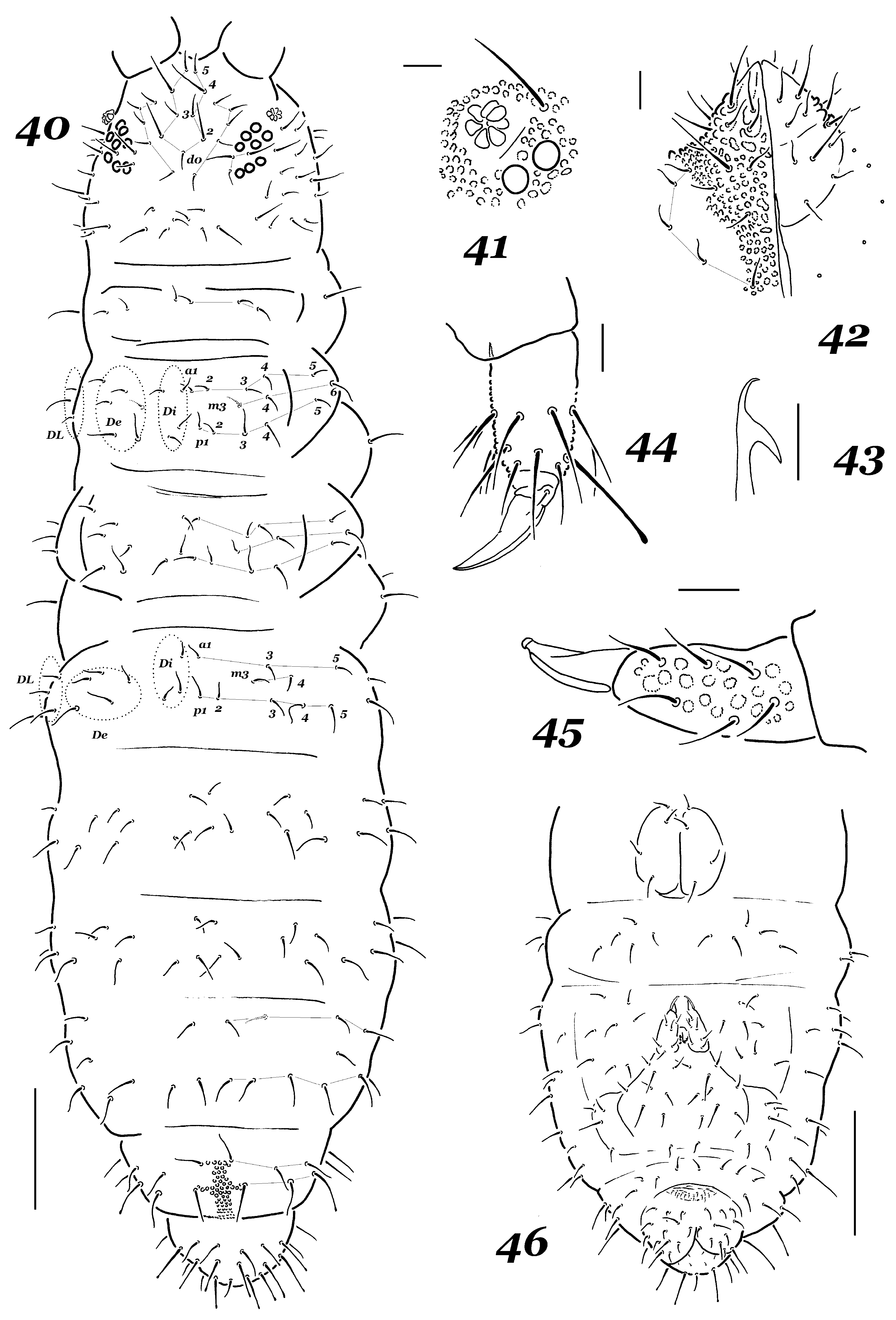
Figs 40–46 View FIGURES 40–46 , Table 1
Type material. Japan: holotype, female, Honshu Island, Nagano Prefecture, E Chino city, Kitayama, surroundings of Mugikusa Hutte, 36°4.13’N 138°20.3’E, 2280 m alt., mixed forest with old trees ( Betula , Abies ) on steep slope, litter, 11 August 2016. M. Potapov & N. Kuznetsova leg. [ MSPU collection]. Paratypes, 4 males and 2 females, same area, but Abies forest along road, under Abies bark, 12 August 2016. M. Potapov & N. Kuznetsova leg. [3 ind.–– MSPU collection and 3 ind.–– TRPM collection].
Diagnosis. Medium sized species. PAO rounded, clearly larger than ocellus. Buccal cone blunt, only two prelabral setae, i.e. 2/2334, totally, labium with 12 ordinary setae and without seta L or labial organites, 5 setae in perilabial area. Dorsal chaetotaxy rich with 3–4 setae in front of setae p3–p4 on Th. II–III, ordinary setae clearly differentiated in length. Mucro with lateral lamella almost reaching the tip. All legs with distinctly clavate tibiotarsal seta A1. Each anal valve with two hr-setae.
Description. Length (without antennae) 0.8–1.1 mm, holotype— 0.9 mm. Alive colour unknown (only cleared specimens available), but probably dark and uniform. Tegument granulation rather coarse.
Antennae slightly shorter than head, Ant. III–IV fused dorsally, ventral separation well marked. Ant. IV with trilobed apical vesicle, external ms, subapical or and seta i present; sensilla (S1–S4, S7–S8) on dorsal side of Ant. IV clearly differentiated, ventral side with only few sensilliform setae. Antennal organ of Ant. III typical, inner sensilla small, sgv almost subequal to sgd (1.0–1.1: 1), ventral ms present. Ant. I–II with 7 and 12 setae, respectively.
Head with 8+8 subequal ocelli. PAO rounded, consisting of 7–8 vesicles, ratio of its diameter to ocellus B as 1.5–1.7: 1 ( Fig. 41 View FIGURES 40–46 ). Buccal cone short and blunt. Maxilla styliform, lamellae unseen. Mandible with two teeth ( Fig. 43 View FIGURES 40–46 ). Distal edge of labrum not prolong, number of labral setae as follows: 2/2334. Main part of labium with four proximal ordinary setae, seta L and labial organites absent; submentum and mentum with usual set of four setae each, i.e. 4+4 ( Fig. 42 View FIGURES 40–46 ). Perilabial area with 5+5 setae.
Dorsal chaetotaxy rather stable ( Fig. 40 View FIGURES 40–46 ), asymmetrical abnormalities not frequent, setae detectably differentiated in length (d2 and d4 on head especially long), longest setae on abdominal tip sometimes truncate or even slightly widened at apex. Sensilla short: on Th. II p1<p3(S)<p4 and on Abd. V p1: p3(S) = 1: 0.9, their number as usual: 22/11111. Head with d0 and without a0. Th. I usually with 3+3 setae (few specimens with 3+4 setae also observed). Th. II with a2-seta and lateral ms present, dorso-external group with three setae (a3–a4, m4) in front of p3–p4. Th. III without a2 and ms as usual but with four a–m setae in dorso-external group (a3–a4, m3–m4). Abd. I–III with 3 setae (a3, m3–m4) in front of p3–p4, occasionally one of them absent. Abd. IV usually with one lateral seta in m-row. Abd. V without p2 as usual.
Thoracic sterna without setae. Ventral tube with 4+4 setae, no seta on sternum of Abd. I, Abd. II with 5+5 sternal setae, Abd. III with 8–9 such setae ( Fig. 46 View FIGURES 40–46 ). Tenaculum with 3+3 teeth as usual. Furca not especially long, reaching posterior border of Abd. II. Manubrium with (8)9+(8)9 setae on main part, 4–5 setae on each basolateral lobe and 2 basal setae in line ( Fig. 46 View FIGURES 40–46 ). Dorsal side of dens with six setae and rather coarse granulation ( Fig. 45 View FIGURES 40–46 ), hyaline field on its ventral side about as large as mucro length. Mucro with upturned tip and long lateral lamella. Each anal valve with two tiny hr-setae.
Legs I–III with usual number of setae: 1, 2, 2 setae on upper subcoxae, 0, 2, 2 setae on lower subcoxae, 3, 7, 7–8 setae on coxae, 6, 6, 6 on trochanters, 13, 12, 11 setae on femora and 19, 19, 18 setae on tibiotarsi, of which A1 elongate and clearly clavate at apex ( Fig. 44 View FIGURES 40–46 ). Unguis with small tooth in mid part of inner edge, lateral teeth absent.
Etymology. The name of the new species reflects its general graceful appearance, concinnus in Latin—harmo-nious, slim, graceful.
Affinities. The most characteristic feature of P. concinnus sp. nov. is the elongated and clearly clavate tibiotarsal seta А1. Among the species recorded in the eastern parts of the Asian continent, only P. dalensi have such setae on tibiotarsi, although their number is much higher (4–4–3 on legs I–III, respectively). The latter species also has some blunt setae on the abdominal tip, two teeth on mandibles, a labium with four proximal setae and without L or organites, PAO with a small number of vesicles, etc. However, dorsal chaetotaxy in P. dalensi is apparently much less complete. In particular, judging by fig. 14C in Weiner & Najt (1985), only 2+2 setae are present on Th. I (3+3 or 3+ 4 in P. concinnus sp. nov.), Th. II without a2 (present in P. concinnus sp. nov.), Abd. I–III with utmost one seta in front of p3–p4 (3 setae in P. concinnus sp. nov.) and Abd. V without a2 (present in P. concinnus sp. nov.).
Perhaps, P. concinnus sp. nov. is even more similar to P. boerneri Sch ӧtt, 1902 , which was also found in the region (see below). Both species have almost identical dorsal chaetotaxy, but are easily distinguishable by the number of clavate tenent setae on tibiotarsi (1,1,1 in P. concinnus sp. nov. vs 6–7,7,7 in P. boerneri ) and differentiation of dorsal setae, which may be truncate or slightly widened on the abdominal tip of P. concinnus sp. nov., but never are knobbed as in P. boerneri .
At least two similar species, P. corticicolus ( Schäffer, 1896) and P. janstachi Kaprus’ & Weiner, 2009, are known from Europe. Both of them also have one clavate seta on each tibiotarsi, PAO with few vesicles and slightly modified dorsal setae on abdominal tip, but can be distinguished from P. concinnus sp. nov. by less complete dorsal chaetotaxy and different labral formulas (4/ 234 in P. corticicolus and 2/ 234 in P. janstachi vs 2/ 2334 in P. concinnus sp. nov.). Apart from these, mandibles appear to differ (with four strong teeth in corticicolus , two in P. concinnus sp. nov. and three in P. janstachi , although a medial tooth in the latter species is tiny and can easily be overlooked), only P. janstachi has a seta L and organites on labium and P. concinnus sp. nov. appears to be unique due to the different lengths of setae on the head.
A fairly similar species, P. columbicus Rusek, 1991 , is known from the other side of the Pacific Ocean. It can be easily distinguished from P. concinnus sp. nov. by the labral formula (4/ 334 in P. columbicus vs 2/ 2334 in P. concinnus sp. nov.), mandibles (three large teeth in P. columbicus vs two teeth in P. concinnus sp. nov.) and peculiarities of the chaetotaxy. In particular, P. columbicus is characterized by the presence of a0 on head (absent in P. concinnus sp. nov.), Th. I with 4+4 setae (3+3 setae in P. concinnus sp. nov.), Th. II–III without setae m3 (always present in P. concinnus sp. nov., at least on Th. III) and by the presence of only two setae in front of p3–p4 on Abd. I–III (three such setae in P. concinnus sp. nov., as a rule).
In North America, a number of other species having clavate tibiotarsal setae are noted. Unfortunately, their morphology is not always fully described, which makes them difficult to compare with P. concinnus sp. nov. Some of them are easily distinguishable, others are obscure, and, for example, « the forms gathered under this name [ corticicolus of American authors] probably represent a cluster of species » ( Christiansen & Bellinger 1980).
Distribution. Known only from the type locality.
| TRPM |
Tottori Prefectural Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |