
Eclipidrilus macphersonae, Fend, Steven V. & Lenat, David R., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.210008 |
|
DOI |
https://doi.org/10.5281/zenodo.6165720 |
|
persistent identifier |
https://treatment.plazi.org/id/BC6B87A2-FFDD-7635-FF5B-FD962C73FEA8 |
|
treatment provided by |
Plazi |
|
scientific name |
Eclipidrilus macphersonae |
| status |
sp. nov. |
Eclipidrilus macphersonae n. sp.
( Figures 5–6 View FIGURE 5 View FIGURE 6 )
Holotype: USNM 1156964: A whole, slide-mounted worm in Canada balsam.
Type locality: North Carolina, Scotland Co., Lumber River near Wagram, 25 March 2011. Collected by D.R. Lenat.
Paratypes: USNM 1156965-66: from the type locality, 25 March 2011, 1 sagittally sectioned worm, 1 whole mount. USNM 1156967: Hoke Co., Flat Creek at Manchester Road, 12 February 2009, 1 whole mount. USNM 1156968-69: Richmond Co., Naked Creek at SR1003, 18 February 2010, 2 whole mounts Collected by D.R. Lenat.
Etymology: Named for Trish Macpherson, in recognition of her contributions to the benthic invertebrate biology of North Carolina.
Other material: North Carolina, Hoke Co., Flat Creek at Manchester Road, 12 February 2009, 3 whole mounts. Richmond Co., Naked Creek at SR1003, 25 January 2010, 1 mature and several immature, whole mounts. 18 February 2010, 4 whole mounts. 6 March 2010, 1 whole mount, 2 sagittally sectioned. Unnamed tributary to Naked Creek at Forest Service road upstream of SR 1003, 10 March 2010, 4 whole mounts. Scotland Co., Lumber River near Wagram, 22 February 2011, 3 whole mounts. 25 March 2011, 3 whole mounts, 1 sagittally sectioned. 14 April 2011, 9 whole mounts. All collected by D.R. Lenat.
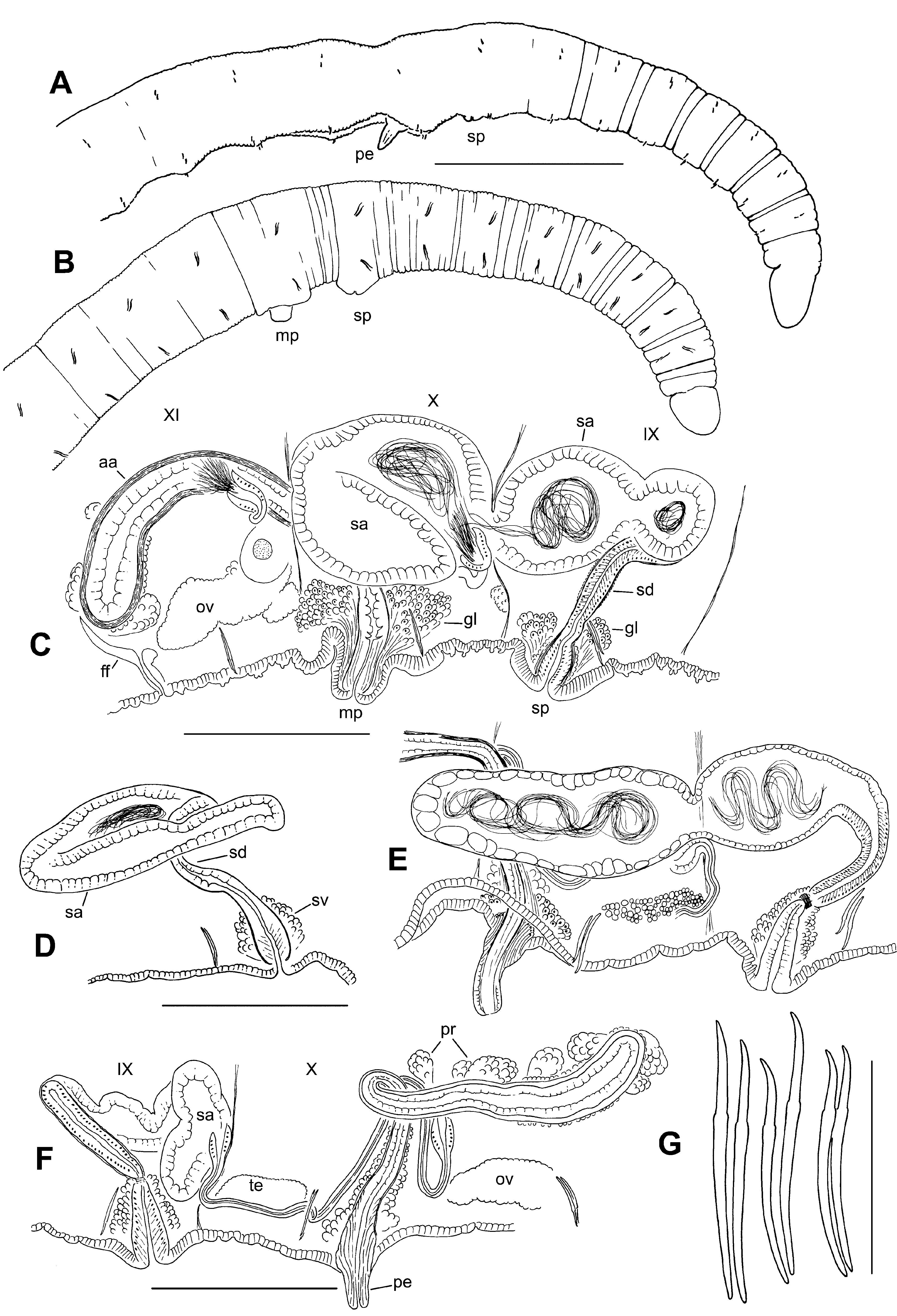
Description. Small, very thin worms, length (preserved) 14–19 mm, 81–107 segments; width 0.19–0.25 mm in X, maximum width to 0.21–0.28 mm. Secondary segmentation a narrow anterior ring in preclitellar segments, from about III–IX ( Fig. 5 View FIGURE 5 A–B). Clitellum indistinct, only slightly thicker or more glandular than surrounding epidermis, (IX)X–XII. Chaetae sigmoid, simple-pointed, with nodulus 31–39 % of chaeta length from tip ( Fig. 5 View FIGURE 5 G). Chaeta length 44–70 μm in mid-body; dorsal and ventral chaetae similar in size; slightly shorter in posterior segments, but proportions similar in anterior and posterior segments. Within each bundle, the outer (more lateral) chaeta may be slightly shorter than the inner. Prostomium rounded to conical, about 0.12–0.15 mm long, width about equal to length; prolobous.
Brain in the peristomium, deeply lobed. Pharynx in II to mid-IV; dorsal wall concave with columnar, ciliated cells, about 12 μm thick; ventral wall very thin (2–4 μm), with cuboidal, non-ciliated cells. Pharyngeal glands well developed dorsolaterally in V–VII or VIII. Longitudinal muscle layer 4–7 μm thick in preclitellar segments; circular muscle layer 2 μm. Septa 1/2 and 2/3 inconspicuous. Epidermis 5–10 μm thick anterior to clitellum; to about 10–12 μm in clitellum; 5–7 μm in post-clitellar segments; to 24 μm in prostomium. Lateral, commissural blood vessels in preclitellar segments; these vessels thin and convoluted, typically joining ventral vessel 1 segment behind junction with dorsal vessel. No lateral blood vessels in middle or posterior segments. Dorsal blood vessel separate in preclitellar segments; adjacent or appressed to top of gut posterior to about X. Chloragogen cells begin in VI or VII.
First nephridia usually paired on 6/7, the next on 12/13; nephridia in a few posterior segments, paired or on one side. Each nephridium has a small anteseptal funnel and a granular postseptal thickening; the posterior duct forms a loop, which extends ventrally, entering one or more posterior segments, and terminating in a short ectal duct in the originating segment. Ectal ducts usually terminate in a nearly spherical vesicle (diameter about 20 μm) at the inconspicuous nephropore.
Spermatheca single, median in IX; pore may be distinct, just behind the ventral chaetae; pore surrounded by a pad of slightly thickened epidermis, 70–120 μm wide, externally appearing as a low mound ( Fig. 5 View FIGURE 5 B). Ectal end of spermathecal duct widens into a narrowly-conical vestibule, 80–120 μm high by 30–45 μm wide near spermathecal pore; the vestibule lined with columnar epithelium, a 4–6 μm thick muscle layer, and surrounded by an irregular (10–22 μm) layer of granular cells ( Fig. 5 View FIGURE 5 C–F, 6D). Narrowed ental end of vestibule joined by a cylindrical spermathecal duct, 110–130 μm long by 22–24 μm wide; duct has columnar epithelium, a very thin muscle layer (about 1 μm) and a narrow lumen; duct narrowed and surrounded by a ring of circular muscle fibers, forming a short sphincter at junction with vestibule ( Fig. 6 View FIGURE 6 D–E). Spermathecal ampulla of fully mature worms (with developed eggs) usually extends into X, sometimes to XI; elongate-sacciform, 360–710 μm long, 60–200 μm wide, containing unordered, loosely-packed sperm ( Fig. 5 View FIGURE 5 C, E). Ampullar epithelium cuboidal in about the ectal 1/5; the remainder thick (12–28 μm), with irregular, vacuolated cells ( Fig. 6 View FIGURE 6 A–C); muscle layer very thin (1 μm). In mated specimens, the sperm is in a loose, unordered bundle ( Fig. 6 View FIGURE 6 C); vacuoles of spermathecal wall may contain sperm ( Fig. 6 View FIGURE 6 B). Spermathecal ampulla of recently-mated worms (without mature eggs) may be folded within IX; with an ovate ectal chamber and a narrower ental portion ( Fig. 5 View FIGURE 5 D); in unmated worms the entire ampulla is narrowly tubular and typically folded ( Fig. 5 View FIGURE 5 F).
Testes paired in IX and X, medium to large size, often extending beyond mid-segment. Ovaries paired in XI, usually large; in some worms extending through XI into XII. Sperm sacs paired, extending back as far as XVII, usually not extending anteriorly from IX; egg sacs may extend 1 or 2 segments beyond sperm sacs. Female funnels 50–95 μm tall, with the posterior side much taller than the anterior; female pores intersegmental, on 11/12.
Male funnels paired on 9/10 and 10/11; simple v-shaped; anterior and posterior pairs about the same size (35– 60 μm high); both directed anteriad, or the posterior may extend back into XI within the sperm sac. Both anterior and posterior male funnels functional, with associated sperm. Both anterior and posterior vasa deferentia very thin (5–8 μm diameter). Posterior vasa deferentia enter X directly, without penetrating 10/11 and forming a loop in XI. All vasa deferentia approach the atrial duct near the beginning of the ampulla, but it is not clear where they enter.
Male pore single, median in X, behind the ventral chaetae ( Fig. 5 View FIGURE 5 A–B); body wall usually concave ventrally in X–XI around and behind pore ( Fig. 5 View FIGURE 5 A, E); concavity not associated with distinct internal musculature ( Fig. 6 View FIGURE 6 G); pore area sometimes protruding as a short, truncate porophore ( Fig. 5 View FIGURE 5 C). Penis cylindrical to truncate-conical, 50– 100 μm long by 22–35 μm wide when extended ( Figs. 5 View FIGURE 5 E, 6F–G), apparently formed by extruded lining cells from the end of the atrial duct, but with small vacuoles visible near the tip. Expanded ectal part of duct (penial structure) about 70–100 μm high, with a 5 μm thick muscle layer, and lined with elongate, outwardly-directed cells having basal nuclei ( Fig. 6 View FIGURE 6 F). Penial structure surrounded by an irregular layer of cells, possibly accessory glands, 17–25 μm thick.
Atrium usually extends into XI or XII. Ectal duct elongate-cylindrical, ascending vertically in X, often forming a loop before junction with the ampulla ( Fig. 5 View FIGURE 5 F). Duct length 130–290 μm, width 16–25 μm, gradually widening entally at ampulla. At the ectal end, duct gradually widens, forming the penial structure. Atrial duct surrounded by a thin (2–5 μm) muscle layer ( Fig. 6 View FIGURE 6 L). Atrial ampulla elongate, cylindrical or club-shaped; length 170–370 μm, maximum width (middle or near ental end) 46–85 μm. Ampullar muscle layer 7–13 μm thick, mostly composed of irregularly transverse fibers, but with a very sparse outer layer of variably oriented diagonal fibers ( Fig. 6 View FIGURE 6 H–K). Ampullar epithelium variable: granular, with indistinct cell boundaries in some specimens, 7–14 μm thick, with basal nuclei ( Fig. 6 View FIGURE 6 H–I); in other specimens thinner and vacuolate ( Fig. 6 View FIGURE 6 J); lumen variable, 5–12 μm wide. Prostate glands multicellular and petiolate, to 30–50 μm tall, bundles generally sparse ( Fig. 5 View FIGURE 5 F, 6I, K); single prostatelike cells may also cover the atria.
Remarks. Although the arrangement of reproductive organs and their general structure bear some resemblance to E. lacustris , the lack of distinct spiral musculature in the atria creates difficulties in attributing E. macphersonae to E. ( Leptodrilus ), or even to the genus. The extrudable penial structures are perhaps more similar to those of some E. ( Eclipidrilus ) than to the E. ( Leptodrilus ) group, and the spermathecal morphology resembles that of E. pacificus ( Fend 2005) . Morphology of the atria, penial structures, and spermathecal ducts is quite similar to that of the western Altmanella species (cf. Fend 2009), but the very different arrangement of these unpaired organs argues against a close phylogenetic relationship. The small size and narrow-elongate body form are distinctive.
Most of the observed E. macphersonae specimens had sperm in the spermathecae, but many of these lacked mature eggs and were probably recently mated. Therefore, the spermathecal ampulla with a broad ectal portion and narrow ental portion seen in these specimens ( Fig. 5 View FIGURE 5 D) appears to represent incomplete development. Expansion of the ental part of the ampulla, with development of the large epithelial cells, must occur well after copulation. Sorption of sperm by these enlarged lining cells has been reported in other Eclipidrilus species ( Fend 2005), as well as in other genera; for example, Fig. 37 in Černosvitov (1930) shows similar vacuoles for Stylodrilus parvus (Hrabë and Černosvitov) , resembling Fig. 6 View FIGURE 6 B in the present paper. One of the specimens having mature eggs in the egg sacs appeared post-mature, with partially resorbed male ducts and altered spermathecal ducts. Nephridia and blood vessels were very difficult to see in most worms, so those observations are based on very few specimens.

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |