
Rhadinella xerophila, Ariano-Sánchez & Campbell, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4442.2.10 |
|
publication LSID |
lsid:zoobank.org:pub:4E2EBF1B-2E73-4FB2-BB7C-1F947EAE7710 |
|
DOI |
https://doi.org/10.5281/zenodo.5975703 |
|
persistent identifier |
https://treatment.plazi.org/id/BB75A160-0010-6068-29F2-FA4100DDFCA8 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhadinella xerophila |
| status |
sp. nov. |
Rhadinella xerophila sp. nov.
Figs. 1–2 View FIGURE 1 View FIGURE 2 , Table 1
Holotype. Adult male, UVG R-7003, collected on 29 October 2016 by D. Ariano-Sánchez in the northern limit of Heloderma Natural Reserve ( HNR) at El Arenal (14.868878 N, 89.790526 W; 580 m above sea level, asl hereafter), Cabañas, Zacapa, Guatemala. The individual was found active crawling on sandy soil in a dry gully at approximately 19:30 h.
Diagnosis. Small, slender and gracile snake of genus Rhadinella , which can be distinguished from the other congeners by the following unique combination of characters: head only slightly distinct from neck; inverted dark U-shaped marking on rostral; anterior supralabials with dark edges and pale centers; conspicuous orange collar on neck divided at mid-dorsum, extending 3.5 scale lengths at mid-dorsum and three scale lengths on side of head, posterior to ultimate supralabial; no subpreocular scale; no broad diastema; temporals 1 + 2; dorsal scale rows 17, smooth, unreduced throughout body; ventrals 158; cloacal scute divided; tail complete, subcaudals 116, divided; dorsal coloration dark gray to blackish, almost totally obfuscating longitudinal striping, ground color not extending to lateral edges of ventrals.
Rhadinella xerophila differs from R. godmani , R. hannsteini (Stuart 1949) , R. hempsteadae , R. kanalchutchan (Mendelson & Kizrian 1995) , R. kinkelini , R. lachrymans (Cope 1970) , R. montecristi (Mertens 1952) , R. pegosalyta (McCranie 2006) , R. rogerromani (Köhler & McCranie 1999) , R. serperaster (Cope 1871) and R. tolpanorum (Holm & Cruz 1994) by having a dark gray dorsal body coloration and absence of conspicuous striping (vs. conspicuous striping and lighter brown color dorsal body coloration). Additionally, the new species differs from R. godmani , R. hempsteadae , R. kanalchutchan , R. montecristi , R. pegosalyta , and R. serperaster by having 17 dorsal scale rows (vs.> 17 dorsal scale rows). Rhadinella xerophila can be most easily confused with R. donaji ( Campbell 2015) , R. dysmica ( Campillo, Dávila-Galavíz, Flores-Villela & Campbell 2016) , R. schistosa (Smith 1941) , R. pilonaorum , and R. posadasi (Slevin 1936) , all of which have almost uniformly dark brown or black dorsum, pale dashes on most dorsal scales, and often an indistinct dark vertebral line. Rhadinella xerophila has a mostly dark head with broad orange edging on supracephalic scales differing from R. donaji with a mostly white head, R. dysmica , R. posadasi , and R. schistosa with mostly or uniformly black heads, and R. pilonaorum with top of head mostly orange with black mottling; differing from R. donaji and R. dysmica by having fourth and fifth infralabials contacting posterior genials (vs. third and fourth); differing from R. donaji , R. dysmica , R. pilonaorum and R. posadasi by having 1+1 temporals (vs. 1 + 2); segmental count differences difficult to evaluate owing to paucity of material, but R. xerophila appears to have more ventrals in males, 158, than do R. schistosa (145–147), R. posadasi (136–141), and R. pilonaorum (151–153), and less than R. donaji (158 vs. 166); number of subcaudals in R. xerophila exceeds the number known for all other dark Rhadinella ( Table 1).
Description of the holotype. Adult male; head length 8.3; SVL 207; TAL 128, eye diameter 0.9; distance of naris to anterior edge of eye 1.6; internasal suture 2.5, about 85% length of prefrontal suture; frontal 2.7, 61% as long as parietals; interocular distance 3.2, 73% occupied by frontal, mental broad, 1.6 x 0.9; two pairs of genials, subequal in size, posterior pair wing-shaped, deeply incised along median suture. Dorsal scales smooth, in 17/17/ 17 rows; apical pits absent; head scales arranged in typical colubrid fashion with paired internasals, paired prefrontals, broad median frontal, narrow paired supraoculars, and paired parietals; supralabials 7/7, fourth and fifth entering orbit; infralabials 7/7; preoculars 1/1, subpreocular absent; postoculars 1/1; temporals 1+1; preventrals 3; ventrals 158; cloacal plate divided; tail complete, subcaudals 116, divided.
Coloration in life. Dorsal ground color dark gray with only a hint of blackish longitudinal stripes on body; center of scales of first two scale rows with paler centers forming somewhat indistinct pale longitudinal lines; top of head black with most supracephalic plates broadly edged with orange; loreal, preoculars, upper and most of lower temporals mottled with orange; supralabials 1–5 and anterior portion of 6 white with blackish posterior edges; nuchal collar orange dorsally and laterally to level just above rictus, ventrally becoming white and merging with ventral coloration; nuchal collar reaching posterior of parietals, mid-dorsally extending posteriorly four scale lengths, laterally extending three scale lengths; nuchal collar partially divided mid-dorsally by several black dashes; chin and gular region whitish, grading to gray, darker posteriorly; iris reddish bronze.
Color after preservation. Dorsum dark brownish gray; top of head dark brown with the orange-auburn markings in life fading to pale brown; nuchal white; and venter mostly cream.
Hemipenial morphology. A complete eversion of the left hemipenis was not achieved. On the basis of partially eversion the organ appears to be simple, capitate with calyculate capitulum; mid-section of hemipenial body with 6–7 large curved spines on sulcate side, asulcate side and base of organ with smaller spines and tiny spinules. The right and left retracted hemipenes extending to ninth and tenth subcaudals, respectively. The sulcus spermaticus is centrolineal, bifurcating just below the capitulum.
Etymology. The specific name is a feminine noun in apposition from the Greek ξηρός (xeros), meaning dry and the Latin phila, meaning loving, in reference to its characteristic of being the sole known member of Rhadinella that inhabits in forest experiencing the harsh and extended dry season characterizing the Motagua Valley. It also represents the urgent need that people and governments take the dry forest seriously as a biodiversity relevant forest that deserves the same levels of protection and love as other more known and charismatic forests such as rainforest.
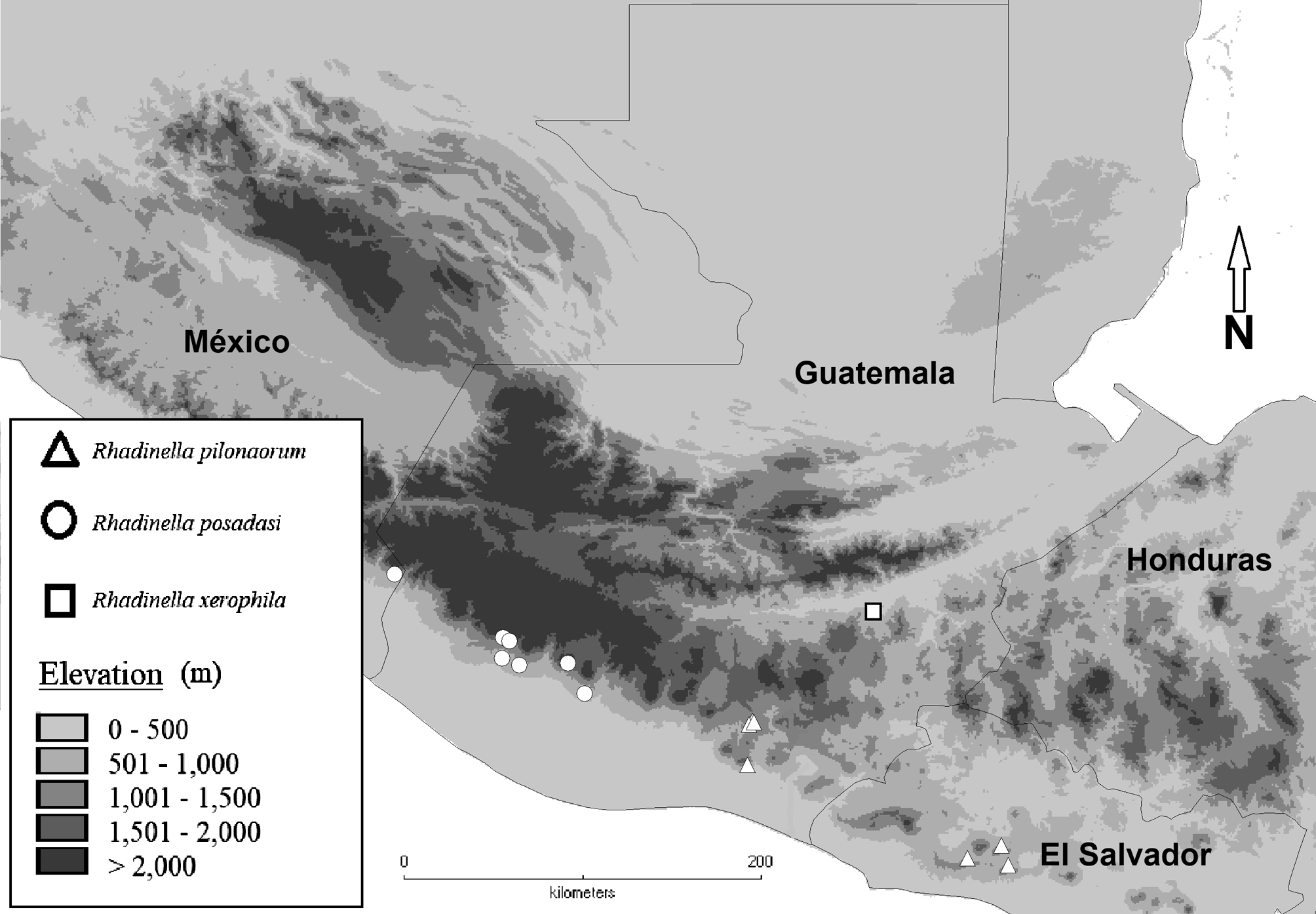
Distribution and habitat. The type locality is located within Heloderma Natural Reserve, a privately owned, protected area within Motagua Valley in eastern Guatemala ( Fig. 3 View FIGURE 3 ). The annual average rainfall (800 mm) in this region is typical of seasonally dry tropical forests. Rainfall in this area is seasonal, with a wet season lasting five months, from June to October, and a dry season of seven months, from November to May. The landscape is composed of patches of seasonally dry tropical forest and tropical thorn scrub, within a matrix of cornfields and melon crops. Most of the topography is undulating, with steep slopes ( Fig. 1B View FIGURE 1 ). The vegetation includes such distinctive dry forest species as Bursera excelsa (Burseraceae) , Nopalea guatemalensis (Cactaceae) , Pilosocereus leucocephalus (Cactaceae) , Stenocereus pruinosus (Cactaceae) , Leucaena collinsii (Mimosaceae) , Lysiloma divaricatum (Mimosaceae) , Bucida macrostachya (Combretaceae) , and Bonellia macrocarpa (Theophrastaceae) . The elevation in this area ranges from 300 to 700 m asl.
Other Motagua valley endemic species of amphibian and reptiles recorded in the vicinity of the type locality are: Oedipina motaguae ( Brodie, Acevedo & Campbell 2012) , Craugastor inachus (Campbell & Savage 2000) , Ctenosaura palearis (Stejneger 1899) and Heloderma charlesbogerti (Campbell & Vannini 1988) .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Rhadinella xerophila
| Ariano-Sánchez, Daniel & Campbell, Jonathan A. 2018 |
Rhadinella xerophila
| Ariano-Sánchez & Campbell 2018 |
Rhadinella xerophila
| Ariano-Sánchez & Campbell 2018 |
Rhadinella xerophila
| Ariano-Sánchez & Campbell 2018 |
R. xerophila
| Ariano-Sánchez & Campbell 2018 |
R. xerophila
| Ariano-Sánchez & Campbell 2018 |
R. dysmica ( Campillo, Dávila-Galavíz, Flores-Villela & Campbell 2016 )
| Campillo, Davila-Galaviz, Flores-Villela & Campbell 2016 |
R. donaji (
| Campbell 2015 |
R. pegosalyta
| McCranie 2006 |
R. rogerromani (Köhler & McCranie 1999)
| Kohler & McCranie 1999 |
R. kanalchutchan
| Mendelson & Kizrian 1995 |
R. tolpanorum
| Holm & Cruz 1994 |
R. lachrymans
| Cope 1970 |
R. montecristi
| Mertens 1952 |
R. hannsteini
| Stuart 1949 |
R. schistosa
| Smith 1941 |
R. posadasi
| Slevin 1936 |
R. serperaster
| Cope 1871 |