
Simulium clarkei, Stone & Snoddy, Stone & Snoddy
|
publication ID |
https://doi.org/ 10.5281/zenodo.4620278 |
|
persistent identifier |
https://treatment.plazi.org/id/BA768705-5129-AC75-0F3F-1411FD09BECA |
|
treatment provided by |
Plazi |
|
scientific name |
Simulium clarkei |
| status |
|
Female. Wing length 2.93.3 mm. Scutum grayish black, unpatterned; thoracic hair silvery, with golden reflections. Radius with hair dorsobasally. Mandible with 2836 serrations; lacinia with 2225 retrorse teeth. Precoxal bridge incomplete. Mesepimeral tuft, basal fringe, and hair of stem vein, basicosta, and scutellum silvery to pale golden. Legs brown, with silvery hair; pedisulcus deeply incised; calcipala well developed; claws each with basal, thumblike lobe. Sensory vesicle in lateral view occupying about 1/31/2 of palpomere III. Anal lobe in ventral view extensively sclerotized; in lateral view with small, unsclerotized, nipplelike extension. Genital fork with each arm expanded into a large, triangular lateral plate bearing prominent, anteriorly directed apodeme. Spermatheca longer than wide, unpigmented proximally; surface with polygonal pattern.
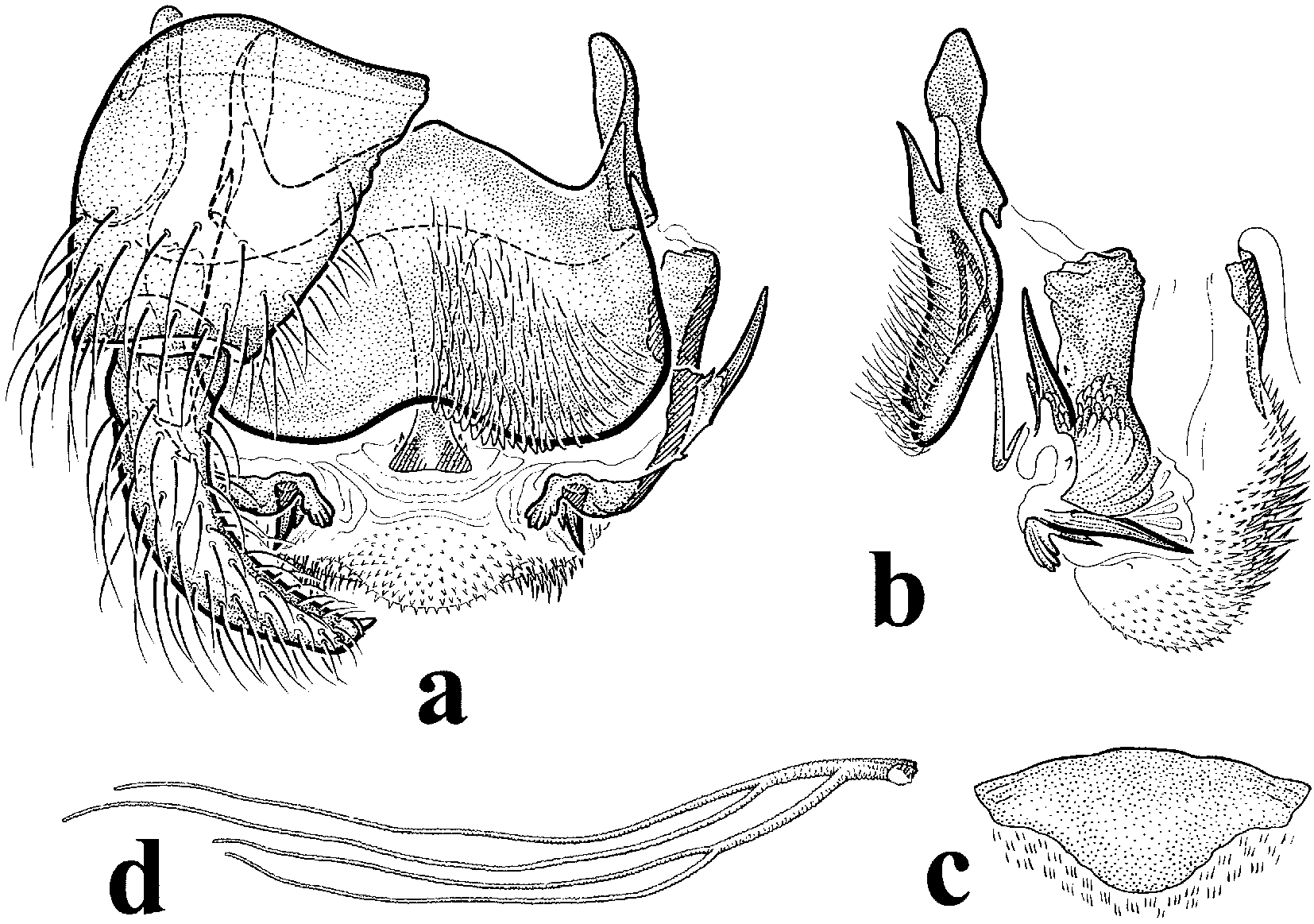
Male. Wing length 2.62.9 mm. Scutum velvety, brownish black; thoracic hair pale golden. Scutellum with coppery to brown hair. Radius with hair dorsobasally. Hair of stem vein, basicosta, mesepimeral tuft, basal fringe, and abdominal hair coppery to brown. Legs brown, with pale golden hair. Genitalia ( Figs. 1 View FIGURE 1 ac): Gonostylus slender, tapered to pointed apex, with 1 apical spinule. Ventral plate in ventral view enormous, well rounded and slightly concave posteriorly; median keel virtually absent. Median sclerite elongate. Dorsal plate subtriangular, wider than long. Paramere with 1 medial and 1 distal spine; aedeagal membrane with numerous smaller spines.
Pupa. Length 3.54.0 mm. Gill ( Fig. 1 View FIGURE 1 d) of 4 filaments, about as long as pupa; base short, giving rise to 2 petiolate pairs of filaments; dorsal petiole about twice as thick and 0.92.0 times as long as ventral petiole; filaments with transverse furrows. Head and thorax dorsally with numerous, irregularly distributed, rounded microtubercles; trichomes simple. Cocoon slipper shaped, without anterordorsal projection.
Larva. Length 6.36.9 mm. Body with reddish brown pigment arranged as spots, or more commonly, as dorsomedially disjunct bands, often densest on first and fifth abdominal segments, ( Figs. 3 View FIGURE 3 a, b). Head capsule pale yellowish brown, contrasting with brown head spots. Antenna pale yellowish brown, extended beyond stalk of labral fan by about 1/23/4 length of distal article; medial article with 4 hyaline bands. Hypostoma with lateral and median teeth small, typically extended anteriorly to about same level. Postgenal cleft straight or biarctate anteriorly, slightly wider than long, extended about 1/3 or less distance to hypostomal groove; subesophageal ganglion with or without ensheathing pigment. Labral fan with 4249 primary rays. Abdominal segment IX with prominent ventral tubercles; abdominal setae simple, pale, sparse. Posterior proleg with 912 hooks in each of 6266 rows. Rectal papillae of 3 compound lobes.
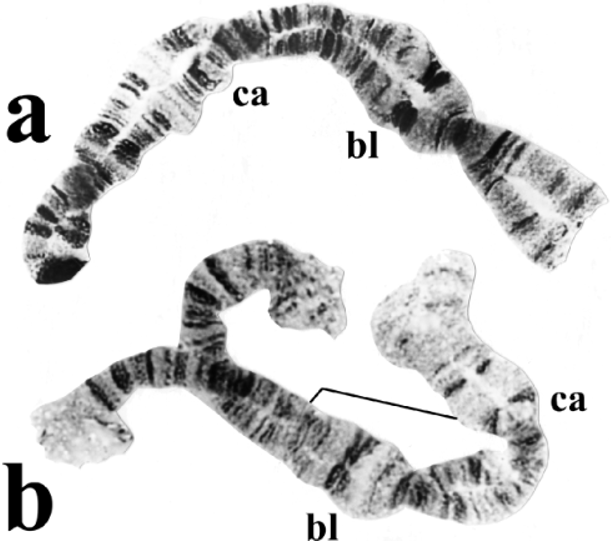
Chromosomes. Larvae have the standard banding sequence of Golini & Rothfels (1984) in chromosome arms IS and IL, and the centromere region of chromosome I is expanded. The tendency to form a chromocenter is negligible, unlike in other members of the group. The following characters are relative to the standard sequence presented by Golini & Rothfels (1984). The IIIS arm is fixed for an inversion (limits 75/7680p; p = proximal) that reverses the ‘blister’ and ‘capsule’ markers. IIL1 is an autosomal inversion, with the inverted sequence predominating in larvae from the Salkehatchie River in South Carolina (3 February 1994) and the standard sequence predominating in larvae from the Meherrin River in Virginia (20 February 2002). IIS is standard in South Carolina, but carries an inversion (limits 49p53p) that reverses the Ring of Balbiani and bulge ‘marker’ in Virginia. This IIS inversion might be indicative of a second species, but in light of the geographic distance between populations in Virginia and South Carolina, we tentatively consider the two populations conspecific. IIIL has an inversion that reverses the orientation of the ‘basal marker’, plus a common floating inversion overtop. Other polymorphisms are common, but sex chromosomes are undifferentiated. The nucleolar organizer is located in the base of IIIL.
Diagnosis. Females are not easily distinguished from those of related species. Males have the largest ventral plate of all North American species. The pupal gill with its four rather smooth, slightly swollen trunks of unequal length — the dorsal trunk being longest — usually permits identification. The brownbanded or spotted body distinguishes the larvae from those of most other North American species with annulated antennae. However, the larvae and pupae can be difficult to distinguish morphologically from those of S. emarginatum Davies, Peterson & Wood , the most similar species, when the two species occupy the same watercourses. Where they occur together, S. clarkei and S. emarginatum can be distinguished by the larval polytene chromosomes and the shape of the male ventral plate. The unique IIIS inversion, which reverses the ‘blister’ and ‘capsule’ markers, provides a powerful diagnostic tool for the larvae of S. clarkei ( Fig. 3 View FIGURE 3 ).
Taxonomic Remarks. Larval pigmentation is positively correlated with water color. Banded larvae are associated with blackwater rivers, whereas spotted to incompletely banded larvae are typical of clearer flows and greenwater rivers ( Fig. 3 View FIGURE 3 ). The petioles of the pupal gill in all specimens in our collections are about half the length of those illustrated by Stone & Snoddy (1969), which are based on specimens from Tennessee, suggesting either that the species is morphologically variable on the two sides of the Appalachian Mountains or that two species are involved. The pupal gill of all material in our collections resembles that of the associated exuviae of the holotype.
Biology. Larvae and pupae of S. clarkei are most common in cold (<12°C), wide, sandy rivers of the piedmont region of the southeastern United States. We also have found them in the upper coastal plain. For example, the Salkehatchie River in the upper coastal plain of South Carolina is teacolored, with a pH between 6 and 7 and a width of 520 m, depending on rainfall. Larvae and pupae are found predominantly on sticks and leaf mats, but sometimes on trailing green vegetation. Simulium clarkei develops primarily in January and February, although stragglers can be found as mature larvae or pupae until mid April.
We have collected S. clarkei with 10 additional species of black flies. However, only one species, Ectemnia primaeva Moulton & Adler , was collected with S. clarkei at more than half the sites, with the Meherrin River collection representing the northernmost record for this species of Ectemnia . The morphologically similar S. emarginatum was collected with S. clarkei at one site. In the Meherrin River, which flows from the Virginia piedmont to the North Carolina coastal plain, our collections suggest that the two species occur in a gradient from pure S. emarginatum to pure S. clarkei .
Distribution. Several trips during the past several years to the type locality of S. clarkei (Blackwater Swamp, Prince George County, Virginia) failed to yield any material of this species, despite intensive efforts in both February and April. In February of 2002, we collected much of the length of the Blackwater River, without finding any specimens of S. clarkei . We eventually found the species during the last week of February in the Meherrin River of North Carolina and Virginia, within 70 km (south) of the type locality. Simulium clarkei is now known from southern Virginia through the Carolinas to southern Georgia. We suspect, based on availability of suitable habitats for the immature stages, that S. clarkei also occurs in Alabama and perhaps as far west as eastern Texas.
Material Examined. GEORGIA, Stewart Co., Pataula Creek, Rt. 27, 31 56.04'N 84 48.12'W, 15 March 1995, P. H. Adler (1 pupa); 18 January 2002, J. K. Moulton (24 larvae, 1 pupa, 4 females & 2 males + exuviae). NORTH CAROLINA, Northampton Co., Meherrin River, Branchs Bridge Road, 36 31.92'N 77 15.68'W, 20 February 2002, P. H. Adler & J. K. Moulton (27 larvae, 3 pupae). SOUTH CAROLINA, Barnwell Co., Salkehatchie River, SSRS6166, 33 17.2'N 81 25.9'W, 3 February 1994, P. H. Adler & J. W. McCreadie (57 larvae, 8 pupae, 7 exuviae, 6 females & 4 males + exuviae); 19 January 2002, J. K. Moulton (63 larvae, 5 pupae, 6 females & 12 males + exuviae). VIRGINIA, Brunswick Co., Meherrin River, Rt. 639, 36 46.26'N 77 58.89'W, 21 February 2002, P. H. Adler & J. K. Moulton (3 larvae).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |