
Gorgosaurus libratus, Lambe, 1914
|
publication ID |
https://doi.org/10.5281/zenodo.3725717 |
|
DOI |
https://doi.org/10.5281/zenodo.3729617 |
|
persistent identifier |
https://treatment.plazi.org/id/B97187EF-FFE0-4A2C-FCE4-FD657CE7FE4C |
|
treatment provided by |
Jeremy |
|
scientific name |
Gorgosaurus libratus |
| status |
|
In the following sections, elements of Gorgosaurus ( Figs. 1–4 View Fig. 1 View Fig. 2 View Fig. 3 View Fig. 4 ) will be described first. The rationale is that this is the most common (and possibly least derived) Alberta tyrannosaurid. The same elements of Albertosaurus are compared ( Figs. 6–16 View Fig. 6 View Fig. 7 View Fig. 8 View Fig. 9 View Fig. 10 View Fig. 11 View Fig. 12 View Fig. 13 View Fig. 14 View Fig. 15 View Fig. 16 ), followed by those of Daspletosaurus ( Figs. 18–36 View Fig. 18 View Fig. 19 View Fig. 20 View Fig. 21 View Fig. 22 View Fig. 23 View Fig. 24 View Fig. 25 View Fig. 26 View Fig. 27 View Fig. 28 View Fig. 29 View Fig. 30 View Fig. 31 View Fig. 32 View Fig. 33 View Fig. 34 View Fig. 35 View Fig. 36 ). The figures of Gorgosaurus , Albertosaurus , and Daspletosaurus are arranged taxonomically, rather than in the order they are referred to in the text. This is to improve the reader’s chances of finding figures when leafing through the paper.
TMP 91.36.500 ( Fig. 1 View Fig. 1 ) is a virtually complete skeleton of Gorgosaurus libratus that is 5.1 m in length from the premaxilla to the tip of the tail is. It was presumably a young adult. Although the neural arches are fused to the centra, the sutures are still visible. Albertosaurus sarcophagus is represented ( Figs. 6–16 View Fig. 6 View Fig. 7 View Fig. 8 View Fig. 9 View Fig. 10 View Fig. 11 View Fig. 12 View Fig. 13 View Fig. 14 View Fig. 15 View Fig. 16 ) by TMP 81.10.1, which was about 8 m long at the time of death, and TMP 86.64.1, an almost complete, 6.5 m long articulated skeleton found near the Tyrrell Museum. Most of the Daspletosaurus specimens are large individuals. However, the best−preserved skull is that of TMP 94.143.1 ( Figs. 18–36 View Fig. 18 View Fig. 19 View Fig. 20 View Fig. 21 View Fig. 22 View Fig. 23 View Fig. 24 View Fig. 25 View Fig. 26 View Fig. 27 View Fig. 28 View Fig. 29 View Fig. 30 View Fig. 31 View Fig. 32 View Fig. 33 View Fig. 34 View Fig. 35 View Fig. 36 ), an animal which was approximately 5.8 m long.
The skull ( Fig. 2 View Fig. 2 ) of TMP 91.36.500 ( G. libratus ) is 640 mm long when measured between the premaxilla and the occipital condyle, and 670mm between the premaxilla and the posteroventral margin of the quadrate. The only smaller, reasonably complete skull of G. libratus (TMP 86.144.1) is disarticulated, but would have been 50cm long. AMNH 5664 (the type specimen of Gorgosaurus sternbergi ) has a slightly larger skull with a lateral length of 678 mm. Cranial fragments and a pair of nearly complete lower jaws (TMP 94.12.155) are from a smaller G. libratus individual whose skull would have been 364 mm long (based on a logarithmic comparison [y = 1.1068x – 0.0317, r 2 = 0.95] of 22 tyrannosaur skulls where both skull and jaw lengths are known).
The small skull of Daspletosaurus sp. (TMP 94.143.1) is about 620mm long between the premaxilla and quadrate.
Small, juvenile skulls are known for two other tyranno−saurids—that of Nanotyrannus lancensis (CM 7541) is 575 mm long (premaxilla to occipital condyle) and that of Shanshanosaurus huoyanshanensis (IVPP V4878) is an estimated 288 mm long ( Currie and Dong 2001).
The skulls of smaller tyrannosaurids are relatively long and low compared to the adults. The maximum cranial width of TMP 91.36.500 is 160 mm across the postorbitals, which is half that of an adult G. libratus (UA 10), suggesting that the skull was relatively narrower at the back in juveniles. The equivalent width measurements in Daspletosaurus sp. (TMP 94.143.1) and Nanotyrannus , which have shorter skulls, are respectively 154 and 210mm. These genera are clearly broader across the postorbital region of the skull at any equivalent age.
The antorbital fenestra of the small G. libratus (TMP 91.36.500) makes up 37% of the antorbital skull length and 62% of the preorbital height. These proportions are almost the same (38%, 63%) in an adult G. libratus (UA 10), which shows that there is little ontogenetic change in the length of the antorbital fenestra during growth.
Unlike allosaurids, sinraptorids and most other theropods, the tyrannosaurid antorbital fossa does not extend onto the nasal. At the level of the back of the maxillary tooth row, the ventral edge of the antorbital fossa extends more than 10 mm below the ventral border of the antorbital fenestra in Gorgosaurus , whereas the two margins almost coincide in Daspletosaurus ( Fig. 16 View Fig. 16 ). A. sarcophagus (NMC 5601, TMP 81.10.1, 85.98.1) seems to have a wider, more horizontal separation between the two margins than G. libratus . The promaxillary slit is relatively smaller in Daspletosaurus ( Fig. 18 View Fig. 18 ) and Tyrannosaurus ( Brochu 2002) than it is in Albertosaurus ( Russell 1970) and Gorgosaurus .
The maxillary fenestra is midway between the anterior margins of the antorbital fossa and antorbital fenestra ( Figs. 2A, B View Fig. 2 , 3A View Fig. 3 ) in Gorgosaurus of any age. Small tyrannosaurine specimens are similar to Gorgosaurus in that the maxillary fenestra is relatively small and central ( Fig. 3E, F View Fig. 3 ). However, mature tyrannosaurines have greatly enlarged maxillary fenestrae with anterior margins that are coincident with the anterior margins of the antorbital fossae ( Fig. 3D, G, H View Fig. 3 ).
The orbits of juvenile tyrannosaurids are only slightly higher than long. Contrary to Carr (1999), this is not so much of a juvenile characteristic as it is of small size, and all theropod adults smaller than tyrannosaurid juveniles also have large, round orbits.
Premaxilla.—The pitted external surface of the premaxilla of Gorgosaurus is higher than long. The supranarial process only diverges slightly from the long subnarial process. The gently curving contact with the maxilla is interrupted by a slit−like subnarial foramen. The distal end of the subnarial process is separated from the maxilla by the nasal. In dromaeosaurids and ornithomimimids, the subnarial process of the premaxilla is wedged between the nasal and maxilla. The subnarial process of the Gorgosaurus premaxilla meets and overrides the nasal beneath the external naris as in Daspletosaurus ( Fig. 18 View Fig. 18 ), Nanotyrannus , Tarbosaurus ( Maleev 1974) , and Tyrannosaurus . This may not be the case in some specimens of Tyrannosaurus ( Brochu 2002) . Russell (1970) reported that the premaxilla did not meet the nasal below the external naris in Daspletosaurus torosus , and used this characteristic to distinguish Daspletosaurus from Albertosaurus . However, the sutures for the premaxilla and nasal on the maxillae (NMC 8506, TMP 89.17.53, 94.172.115) demonstrate that the subnarial processes of Daspletosaurus met below the external naris in all specimens.
The nasal process in immature and mature specimens of Gorgosaurus , Albertosaurus , juvenile Daspletosaurus , and juvenile Tarbosaurus bataar (GIN 100/777) are distally forked. The medial process of the fork sits in a groove on the dorsal surface of the nasal, and the distal ends of the paired premaxillae are separated from each other by the nasals. In Nanotyrannus , Tyrannosaurus ( Holtz 2001, Brochu 2002), and mature specimens of Daspletosaurus and Tarbosaurus , the nasal processes are closely appressed and taper posteriorly to separate the anterior tips of the nasals. This character is gradational with the posterior tips of the nasal processes always being separated in albertosaurines and juvenile specimens of Daspletosaurus and Tarbosaurus . At least some mature Daspletosaurus torosus (NMC 8506) and T. rex specimens (BHI 3033) show a remnant of the groove in the nasal for the posterior tip of the premaxilla. The nasal processes of the premaxillae are separate distally in at least one specimen of T. rex (TMP 81.6.1).
In ventral view, the teeth of the paired premaxillae of G. libratus (TMP 91.36.500) form a semicircle that is wider than anteroposteriorly long. The premaxilla forms the anterior wall of the alveolus for the first maxillary tooth, an unusual arrangement amongst theropods.
Maxilla.—Almost half the maxillary length of G. libratus (TMP 91.36.500) is in front of the antorbital fenestra. One of the foramina on the external surface is relatively large and anteriorly oriented, and is connected to the subnarial foramen by a depression ( Fig. 2A View Fig. 2 ). As in Albertosaurus ( Fig. 6A View Fig. 6 ), Alioramus ( Kurzanov 1976), Daspletosaurus ( Fig. 18A View Fig. 18 ), and Tyrannosaurus ( Brochu 2002) , there is a second major row of foramina that arches above the alveolar margin.
The posterior end of the maxilla bifurcates into two processes ( Fig. 2A View Fig. 2 ), the lower of which underlies the jugal. The medial surface of this process extends higher than the lateral surface, and the jugal sits in a shallow trough between the two. The upper process is a vertical plate that separates two processes at the front of the jugal, the outer one of which covers the lateral surface of the maxilla. The medial edge of the distal end of the maxilla reaches the ectopterygoid ( Fig. 2B View Fig. 2 ), although there are no conspicuous contact surfaces on either bone.
In all tyrannosaurids, the antorbital fossa forms a smoothwalled depression in the ventral margin of the posterodorsal process. Dorsally there is a laterally rugose ridge that separates the antorbital fossa and the nasal other than for a short distance posteriorly. Distally, the posterodorsal process bifurcates to embrace the margins of the anterodorsal process of the lacrimal. The lower of the two prongs contacts the lower surface of this process, whereas the shorter upper prong overlaps the lateral surface. In TMP 83.36.100 ( G. libratus ), the lacrimal extends anteriorly more than 5 cm along the medial surface of the posterodorsal process of the maxilla. The maxilla−lacrimal contacts of A. sarcophagus (TMP 86.64.1) and Daspletosaurus ( Fig. 18A View Fig. 18 ) are similar, although the anterior tip of the lacrimal is not separated from the maxilla by the nasal in mature specimens of Daspletosaurus sp. (TMP 85.62.1, 98.48.1), Tarbosaurus ( Hurum and Sabath 2003) , and some specimens of Tyrannosaurus .
The anterodorsal process on the medial side of the maxilla in tyrannosaurids protrudes only a short distance beyond the end of the bone to contact the inner surface of the premaxilla. The intermaxillary suture of G. libratus (TMP 85.11.3) has three prominent ridges and grooves above the first four maxillary teeth along the internal surface of the palatal shelf of the maxilla. The vomer overlapped the ventral surface at the back of this process. From a point above the fourth maxillary tooth to the level of the anterior margin of the antorbital fenestra above the eighth maxillary tooth, the internal margin of the palatal shelf is smooth and rounded to form part of the boundary of the internal naris. Behind this point there is a well−defined suture for the palatine.
The lower surface of the palatal shelf in Gorgosaurus has pronounced depressions that coincide with the positions of dentary teeth. The four deepest depressions are between maxillary tooth positions 2–3, 3–4, 4–5 and 5–6. These correspond to mandibular teeth 4 to 7, which are the longest teeth of the dentary. In tyrannosaurids preserved with closed jaws, the dentary teeth are found resting in these depressions. A second, less distinct set of depressions is dorsomedial to maxillary tooth positions 9 to 13 in Gorgosaurus , but is much shallower in Tarbosaurus bataar (PIN 551−1) and T. rex (BHI 3033, UCMP 118742). Similar depressions are much more pronounced and can be seen along the entire palatal shelf of A. sarcophagus ( Fig. 6B View Fig. 6 , TMP 95.25.83, TMP 98.63.88, TMP 99.50.140), which seems to be an autapomorphic character.
The bottom of the maxillary fenestra in the small G. libratus (TMP 91.36.500) is positioned a centimetre above a line drawn between the lower margins of the external naris and the antorbital fenestra. In medial view, the maxillary fenestra opens into the maxillary antrum, which in turn is widely open medially. It is bound dorsally in TMP 83.36.100 ( G. libratus ) and other specimens by a low but conspicuous ridge to which attached a sheet of cartilage ( Witmer 1997) or bone (maxillary antrum wall of Brochu 2002). This separation between the nasal passage and the maxillary antrum is osseous in many primitive theropods ( Currie and Zhao 1993a). The postantral pillar is a much thinner version of the posteromedial plate of bone that encloses the maxillary antrum in Allosaurus ( Madsen 1976) , Deinonychus ( Ostrom 1969) , Sinraptor ( Currie and Zhao 1993a) and Troodon ( Currie 1985) . There are relatively high but separate interdental plates ( Fig. 6B View Fig. 6 ) in all tyrannosaurids.
Nasal.—Tyrannosaurid nasals are coossified, although the suture is open anteriorly to a level behind the external nares, and can still be seen posteriorly between the lacrimals. The nasals must have coossified at a very early age in tyrannosaurids because even small nasals of Tarbosaurus and T. rex (LACM 28471) are fused to the same degree. The internasal suture of Eotyrannus also seems to be completely fused ( Hutt et al. 2001). In Gorgosaurus , Albertosaurus ( Fig. 7 View Fig. 7 ), and Eotyrannus , the nasals are broadest immediately behind the external nares, are constricted by a third between the anterior tips of the lacrimals, and expand at the front of the prefrontals. This is similar in juveniles of Daspletosaurus ( Fig. 18B View Fig. 18 ) and Tarbosaurus bataar (GIN 100/66), but the nasals of mature individuals of these two genera and Tyrannosaurus are further constricted behind the junction of the nasal, lacrimal and maxilla. The constriction is extreme in large specimens of T. rex (BHI 3033) where the nasal is only one sixth its maximum width ( Holtz 2001). In cross−section, the surface of the paired nasals is dorsally convex and rugose back to the level of the lacrimals ( Fig. 2A View Fig. 2 ). Behind this point, the rugose surface splits and diverges to become continuous with rugosities on the anterolateral surfaces of the lacrimals, leaving a flat, smooth dorsal surface on the nasals.
Anteriorly, the nasals diverge to clasp the back ends of the premaxillae ( Fig. 2C View Fig. 2 ). In addition, each nasal splits anteriorly to send a short prong to separate the posterior tips of the premaxillae as in mature specimens of G. libratus (TMP 94.12.602), A. sarcophagus (TMP 86.64.1), juvenile Daspletosaurus ( Fig. 18B View Fig. 18 ), and juvenile Tarbosaurus bataar (GIN 100/777). The nasal process of the premaxilla overlaps the nasal between its two anterodorsal prongs. There does not seem to be a medial prong in mature specimens of D. torosus (NMC 8506), Daspletosaurus sp. (TMP 98.48.1) or Tarbosaurus bataar (ZPAL MgD−I/4), or in juveniles of T. rex (LACM 23845, TMP 81.6.1). This is clearly not an easy character to code given ontogenetic changes and preservational distortion.
Beneath the naris, the nasal has tapering process that sits in a shallow groove on top of the maxilla. The subnarial process of the premaxilla overlaps it anteriorly. The nasal forms the dorsal boundary of the antorbital fossa for only a short distance in G. libratus (4 cm in TMP 91.36.500, 2 cm in TMP 83.36.100 and UA 10). There is no pneumatic invasion of the nasal such as is seen in Allosaurus , Monolophosaurus , Sinraptor and other carnosaurs ( Currie and Zhao 1993a).
Along the entire ventrolateral margin, the nasal has a system of tongue and groove contacts for the maxilla, lacrimal and prefrontal. Most of the subnarial process has a rounded edge that sits in a groove on the dorsal surface of the maxilla. Behind the external naris, however, both the nasal and the maxilla have grooves along the edges where they are in contact. The contacts are fundamentally the same in larger, mature specimens of Gorgosaurus and Albertosaurus ( Fig. 6C View Fig. 6 ), and the juvenile specimen of Daspletosaurus ( Fig. 18A View Fig. 18 ). In contrast, transverse ridges break up this groove in mature tyrannosaurines ( Hurum and Sabath 2003). The back of this groove is bound laterally by a fingerlike process that wraps around the anterior tip of the lacrimal. This process is absent in mature specimens of Daspletosaurus , Tarbosaurus , and some specimens of Tyrannosaurus ( Hurum and Sabath 2003) , but is found in the juvenile specimen of Daspletosaurus ( Fig. 18A View Fig. 18 ) and in several specimens of T. rex (TMP 81.6.1, Brochu 2002). Near the back of the nasal is a simple butt joint for the prefrontal. In lateral view, the curvature of the nasal maxillary suture is very uniform in tyrannosaurids. The margin of the nasal is concave behind the subnarial process, but becomes convex above the antorbital fenestra. Overall, the maxillary−nasal contact is an elaborate articulation that allowed some mediolateral rotation of the maxilla in albertosaurines and juvenile tyrannosaurines.
The dorsal surface of the nasal of G. libratus (TMP 91.36.500) is pierced by a row of about ten conspicuous foramina that pass anterodorsally from the inner surface of the bone. The most anterior foramen is at the back of the external naris, and the most posterior is level with the lacrimal “horn”. Additional, more randomly distributed foramina are found throughout the sculptured region, especially at the base of the subnarial process. A similar foramen distribution is evident in Albertosaurus ( Fig. 6A View Fig. 6 ) and Daspletosaurus ( Fig. 18B View Fig. 18 ). There are fewer of these foramina in Tarbosaurus and Tyrannosaurus ( Hurum and Sabath 2003) .
In Gorgosaurus , the most posterior point of the nasal is at its posterolateral corner ( Fig. 2C View Fig. 2 , NMC 2120, TMP 94.12.602, TMP 2000.12.11, USNM 12814), which is also true for some specimens of Daspletosaurus sp. ( Fig. 18B View Fig. 18 , TMP 98.48.1), Nanotyrannus , Tarbosaurus bataar (PIN 553−1), and T. rex (AMNH 5027, BHI 3033, FMNH PR2081, LACM 23845, TMP 81.6.1). In contrast, the midline process of the nasal extends as far or farther posteriorly than the posterolateral corner in Albertosaurus ( Fig. 6A View Fig. 6 ) and some specimens of Daspletosaurus (NMC 8506, TMP 85.62.1) and Tyrannosaurus . Because of the variability in Daspletosaurus and Tyrannosaurus , this character ( Holtz 2001) may have questionable use in phylogenetic analyses.
In dorsal aspect the posterior margin of the nasal turns anteromedially at an acute angle from the lateral margin. This part of the nasal overlaps a long tongue−like process of the frontal. However, near the midline there is always a short but conspicuous posterior process (the medial frontal process of Carr 1999) in tyrannosaurids that along with its mate from the other nasal sits in a conspicuous midline groove in the dorsal surface of the paired frontals. In TMP 91.36.500 ( G. libratus ), this process is more than 2 cm long. A groove in the dorsal surface of the nasal separates the base of the process from the more lateral region of the nasal.
As pointed out by Russell (1970), the paired nasals expand posteriorly between the lacrimals in Gorgosaurus ( Fig. 2C View Fig. 2 ) and Albertosaurus ( Fig. 6A View Fig. 6 ). This is presumably the plesiomorphic state because it is also found in carnosaurs like Sinraptor ( Currie and Zhao 1993) , and in the primitive tyrannosauroid Eotyrannus ( Hutt et al. 2001). The nasals remain more or less the same width in this region in small specimens of Daspletosaurus ( Fig. 18B View Fig. 18 ) and Tarbosaurus bataar (GIN 100/66) or become narrow between the anterior tips of the prefrontals in larger specimens. The nasals also narrow posteriorly in Tarbosaurus and T. rex (LACM 23845), although it is not as conspicuous in Nanotyrannus (determined by the width of the nasal suture on the frontal) and juvenile specimens of T. rex (LACM 28471). The nature of the contact between the front of the lacrimal and the nasal is also characteristic of certain genera. Like G. libratus and A. sarcophagus , each of the nasals of several T. rex (BHI 3033, LACM 23845, TMP 81.6.1) have a distinct finger−like, lateral process that clasps the anterior end of the lacrimal. This is not evident in full−grown Daspletosaurus sp. (TMP 85.62.1, 98.48.1) and T. rex specimens (LACM 23844). Reduction of this process has previously been identified as an ontogenetic trend ( Carr 1999).
Lacrimal.—In Gorgosaurus ( Fig. 2A View Fig. 2 ) and Albertosaurus ( Fig. 6B View Fig. 6 ), the dorsolateral margin of the lacrimal is raised into a ridge that forms a conspicuous lacrimal “horn” anterodorsal to the orbit. Ontogenetically, it is poorly developed in juvenile G. libratus (TMP 86.144.1), is sharp and pronounced in young adults ( Fig. 2A View Fig. 2 ), and becomes wider, more massive and less pronounced in large individuals (NMC 2120, TMP 94.12.602). Similar trends are evident in A. sarcophagus ( Fig. 6B View Fig. 6 , TMP 81.10.1, 86.64.1). Juvenile Daspletosaurus ( Fig 18A View Fig. 18 ) have a low mound in the same position, but as in other tyrannosaurines this develops into an elongate, inflated ridge in adults (NMC 8506, TMP 85.62.1, 2001.36.1). MOR 590, a possible new species of Daspletosaurus , is intermediate in size and inflation of the lacrimal. An immature specimen of Tarbosaurus bataar (GIN 100/66) has pronounced lacrimal horns that are more conspicuous in lateral view than the postorbital boss. In large specimens of tyrannosaurids, the interiors of the lacrimal “horn” (in albertosaurines) and swelling (in tyrannosaurines) are hollow. The bone surrounding the sinus can be less than a few millimeters thick, and in large individuals like MOR 555 is penetrated by what appear to be small pneumatic foramina. It is unlikely that tyrannosaurs engaged in head butting ( Bakker 1986, Bakker et al. 1988), even though the frontals and parietals are disproportionately thick, solid bones. Carr (1999) referred to the lack of a horn in Tyrannosaurus and Tarbosaurus , although this apparent lack is due to the inflation of the entire dorsal ramus of the lacrimal into an elongate ridge rather than a conspicuous horn. The thickness of the lacrimals in these genera and Daspletosaurus are comparable to the height of the horn in Gorgosaurus . For example, the lacrimal horn is 57 mm high in TMP 94.12.602 (a large G. libratus ), but is 70mm high in a slightly larger Daspletosaurus sp. (TMP 85.62.1).
The center of the horn is partially size dependent. In small specimens of G. libratus like TMP 91.36.500, it is positioned over the pneumatic fossa ( Fig 2A View Fig. 2 ). In larger specimens of G. libratus (UA 10), the center is above the front of the pneumatic fossa as in small specimens of Daspletosaurus ( Fig. 18A View Fig. 18 ) and large specimens of Albertosaurus ( Fig. 6B View Fig. 6 ).
In dorsal aspect, the lacrimal is a curved plate of bone with limited exposure ( Figs. 2C View Fig. 2 , 6A View Fig , 18B View Fig ). A millimeter wide slot in the small G. libratus (TMP 91.36.500) that gives the frontal access to the orbital rim separates the lacrimal from the postorbital. This gap closes in the largest specimens of G. libratus (TMP 94.12.602, UA 10) as the lacrimal contacts the postorbital.
Anteriorly, the lacrimal of Gorgosaurus sits in a groove on top of a long tapering process of the maxilla, and bifurcates at its contact with the maxilla and nasal. The lower process extends along the medial surface of the posterodorsal process of the maxilla, whereas the tapering upper process wraps around onto the dorsal surface of the nasal to sit in a shallow groove medial to the fingerlike process.
Posterodorsally, the tyrannosaurid lacrimal is separated from the back of the nasal and front of the frontal by the prefrontal. The crescentic prefrontal sutures on the lacrimals of an immature A. sarcophagus (TMP 86.144.1) extend part way down the preorbital bar. The posterior end of the lacrimal of most tyrannosaurids plugged into a concave, vertically oriented suture on the frontal.
In lateral view, the preorbital bars of tyrannosaurid lacrimals are convex anteriorly and concave posteriorly. At least two lacrimal ducts penetrate the medial ridge on the preorbital bar at midheight. The preorbital bar is strengthened by a vertical ridge ( Fig. 19 View Fig ) on the medial surface, which gives it a T−shaped cross−section at midheight. Dorsally the ridge is continuous with the ventral process of the prefrontal. Ventrally the ridge twists anteriorly to wrap around the back of the anterodorsal process of the jugal. It extends beyond this region to gain lateral exposure, although it is still separated from the lateral surface of the preorbital bar by a conspicuous vertical groove (within the base of which the anterodorsal process of the jugal sits). The two laterally exposed plates diverge ventrally as the smooth−surfaced, more medial one expands anteriorly. It passes onto the inside surface of the jugal where it has a weak, squamose suture that does not contact either the maxilla or the ectopterygoid.
The lateral surface of the lacrimal passes to the outside surface of the jugal where it forms a squamose suture that is triangular in outline. In Gorgosaurus and Albertosaurus , this ventrolateral process forms the posterior margin of the antorbital fossa as it leads into the pneumatopore in the jugal. As pointed out by Russell (1970), this is not the case in Daspletosaurus , where the ventrolateral process does not form a conspicuous ridge. Nanotyrannus , Tarbosaurus and Tyrannosaurus are like Daspletosaurus in this feature, so it appears to be a good character for identifying tyrannosaurines. This contact is strengthened posteriorly by a thick, fingerlike process of the jugal that extends at least 1.5 cm along the back of the lacrimal. In tyrannosaurines, the posteroventral corner of the lacrimal plugs into a socket that is supported by a small but distinct ridge on the external surface of the jugal.
In the posterodorsal corner of the antorbital fossa is found the large pneumatic opening. The triangular opening in the small G. libratus (TMP 91.36.500) is separated from a smaller, more anterior pneumatopore by a vertical bar of bone. Although the shapes and sizes of these openings vary considerably in different specimens, this is a characteristic arrangement in all specimens of Gorgosaurus . In larger specimens, the two pits are sunk into the floor of a larger depression, formed when a new vertical ridge of bone develops anteriorly ( Carr 1999). The pneumatic opening of Daspletosaurus (TMP 85.62.1, TMP 83.30.1) is relatively small and round compared to those of Gorgosaurus and Albertosaurus . Again, this can also be attributed in part to size, and also can be correlated with ontogenetic changes in size. In juvenile Daspletosaurus ( Fig. 18A View Fig ) and Tarbosaurus bataar (GIN 100/66), the pneumatic openings of the lacrimals are relatively larger than they are in the adults. There are nevertheless significant differences in the pneumatization of the lacrimal in different tyrannosaurids. In Albertosaurus and Gorgosaurus , the bridge of bone separating accessory openings from the main pneumatopore is relatively short anteroposteriorly. In Daspletosaurus , the bridge is larger than the size of the main pneumatopore. Tarbosaurus bataar (GIN 100/66, GIN 107/2, PIN 551−1) does not have accessory openings.
The anterodorsal process of the lacrimal extends to about the middle of the antorbital fenestra in Gorgosaurus ( Fig. 2A View Fig ), Albertosaurus (5B), and juveniles of Daspletosaurus ( Fig. 18A View Fig ). Anteriorly, the upper long, slender and tapering process of the lacrimal in Daspletosaurus sp. (TMP 85.62.1) extends anteriorly far beyond the more ventral process to separate the maxilla and nasal for almost the entire length of the antorbital fenestra (in Gorgosaurus it extends less than half this distance). In consequence, the lacrimal is longer than it is tall. Furthermore, mature specimens of Daspletosaurus sp. (TMP 85.62.1, 98.48.1) lack the finger−like process on the nasal that clasps the anterior tip of the lacrimal.
Prefrontal.—The prefrontal is always exposed on the skull roof of tyrannosaurids, although the degree of exposure varies. Its presence is often not detected because it is small, sometimes is overridden by adjacent bones during postmortem distortion of the skull, and in some specimens it has fused indistinguishably to one of the adjacent bones. The prefrontal of the small G. libratus TMP 91.36.500 is long (61.5 mm) and narrow (12 mm). In one large G. libratus (UA 10), the prefrontal is distinct but seems to be fused onto the anterolateral process of the frontal, whereas in another specimen (TMP 94.12.602) it shows stronger association with the lacrimal. The anterior end of the prefrontal of Gorgosaurus is only slightly anterior to the anterior limit of the dorsal exposure of the frontal. The thin, tapering ventral process ends just dorsal to the lacrimal ducts. The prefrontal tends to be more elongate in Daspletosaurus , where it is exposed on the skull roof anterior to the anterior tip of the frontal (NMC 8506, TMP 85.62.1). It is exposed on the skull roof of Tarbosaurus bataar ( Maleev 1974, GIN 107/2) and some specimens of T. rex (AMNH 5027, TMP 81.6.1).
Frontal.—The frontal of the immature G. libratus (TMP 91.36.500) appears relatively long and narrow (the width to length ratio is 0.43, compared with 0.66 in UA 10). The anterior end of the frontal is overlapped on the midline by a tapering prong of the nasal, and more laterally by a second, longer process. In A. sarcophagus (TMP 81.9.1, 81.10.1, 85.98.1), the nasal−frontal suture is more complex, the midline slot for the nasal expands rather than tapers and extends farther posteriorly than any of the more lateral ridges and grooves for the nasal. The prefrontal and lacrimal have more vertical contacts with the frontal in Gorgosaurus , the latter plugging into a socket (TMP 67.14.3, 80.16.485, 81.39.8, 82.16.181, 91.36.500) in the anterior face of the frontal as in Daspletosaurus (NMC 11841, TMP 80.16.924, 94.143.1). In contrast, the prefrontal seems to have more limited dorsal exposure in A. sarcophagus (TMP 81.9.1, 81.10.1) and the lacrimal did not plug into a socket in the frontal, which is similar to T. rex (RSM 283.2, LACM 23845). However, in the AMNH braincase, there is a deep pit for the lacrimal and the prefrontal seems to have been the same as in Daspletosaurus . Furthermore, in an immature T. rex (LACM 28471), the pit is shallow posteriorly but deep medially. The anterior part of the suture with the postorbital is tall and nearly vertical, whereas posteriorly (within the boundaries of the supratemporal fossa) the postorbital overlaps a shelf on the frontal. The distinction between these two sutural surfaces is less obvious in Daspletosaurus and Nanotyrannus . As a direct consequence, the anterior margin of the supratemporal fossa tends to be more distinct in Gorgosaurus than it is in Daspletosaurus and Nanotyrannus .
The parietal wraps around the posterior margin of the frontal, and sends a small tapering process forward that overlaps the frontal on the midline. The dorsal margin of the frontal rises slightly to meet this process, and a foramen passes posteroventrally through the junction of the two frontals and the overlapping medial process of the fused parietals. In Alioramus ( Kurzanov 1976), Daspletosaurus sp. ( Fig. 20B View Fig , FMNH PR308, TMP 85.62.1), Nanotyrannus , Tarbosaurus , and Tyrannosaurus , the posterior quarter of the frontals rises to take part in an elevated sagittal crest that is much more pronounced than it is in Albertosaurus and Gorgosaurus . It rises sharply above the dorsal surface of the frontal anterior to the supratemporal fossa. For example, TMP 91.36.500 (an immature G. libratus ) represents a larger animal than TMP 94.143.1 (an immature Daspletosaurus ) but the frontal of the latter rises more than double the height at the sagittal crest. Because of these high sagittal ridges, the foramen that is at the junction of the frontals and parietals in G. libratus and A. sarcophagus is roofed over by the frontals and opens in a more anterior position in tyrannosaurines.
The frontal undergoes dramatic ontogenetic changes. Using measurements for the frontals of five tyrannosaurid species ( Table 2 View Table 2 ), the coefficients of allometry were calculated for frontal length and frontal thickness. The correlation coefficient was relatively low for length (r 2 = 0.70), mostly because of variable preservation. Nevertheless, comparison of the logarithms of frontal width versus length shows strong negative allometry (k = 0.48). On the other hand, the thickness (depth) of the bone compared to the width undergoes strong positive allometry (k = 1.38, r 2 = 0.94), probably to form a solid anchor for the jaw musculature. Frontal length is correlated with the length of the brain, which becomes relatively smaller in larger animals. The width of the skull roof increases dramatically during growth, growing outwards far beyond the orbital slots.
Parietal.—The parietal of G. libratus (TMP 91.36.500) has the characteristic tyrannosaurid sagittal crest, extending from the junction with the frontals to the nuchal crest. It is low anteriorly, and in lateral aspect dips posteriorly to a level below the interorbital region of the frontals. There it forms a high knife−like ridge between the supratemporal fenestrae. Posteriorly the sagittal crest rises to the level of the interorbital region of the frontals again. This arrangement is the same in all well−preserved specimens of Gorgosaurus and A. sarcophagus (TMP 81.9.1, 81.10.1), other than UA 10 where the lowest point on the sagittal crest is slightly higher than the interorbital region of the frontals. The sagittal crests on the parietals of Daspletosaurus sp. ( Fig. 21 View Fig , FMNH PR 308, NMC 11841, TMP 80.16.924, 85.62.1), Nanotyrannus and Tyrannosaurus are relatively higher than those of Albertosaurus and Gorgosaurus , mostly by virtue of the fact that the sagittal crest is much higher at the frontoparietal suture of these animals. This gives the crest of Daspletosaurus a more knife−like appearance, whereas the crest seems to rise higher at the nuchal crest in Gorgosaurus and Albertosaurus .
The fused parietals have an anterior process that inserts between the frontals on the midline. Frontals and parietals are elevated along their contact, and in more mature tyrannosaurid individuals (TMP 67.14.3, 81.9.1, 81.10.1) the suture is strengthened by small interdigitating processes. The parietal extends as a long tapering process along the dorsal margins of the exoccipital−opisthotic unit. Much of this process is covered by the squamosal. The parietal forms most of the characteristic tyrannosaurid nuchal crest. The nuchal crests of Gorgosaurus and Albertosaurus are significantly higher but narrower than those of equivalent sized specimens of Daspletosaurus ( Fig. 21 View Fig ) or Nanotyrannus . This correlates with the narrower temporal fenestrae of Gorgosaurus and Albertosaurus .
Postorbital.—In large individuals of Gorgosaurus , Albertosaurus , Daspletosaurus , Tarbosaurus , and Tyrannosaurus , the postorbital establishes contact with the lacrimal.
In immature G. libratus (TMP 91.36.500), a relatively small, rugose tuberosity rises above the lateral surface of the postorbital along the orbital margin. The size of the tuberosity is dependent on the age and size of the individual, and it is not evident in smaller specimens (TMP 86.144.1, G. libratus ). However, regardless of size, the tuberosity is always positioned at the edge of the bone between the dorsal and posterior margins of the orbit. The postorbital is sculptured along the margin of the orbit anterodorsal to the tuberosity, but it is smooth along the edge of the upper temporal fenestra in Gorgosaurus . In large individuals of Albertosaurus ( Fig. 8A View Fig ), the rugose region extends along the entire upper edge of the postorbital anterior to the squamosal, and the tuberosity is more prominent. The size of the postorbital “horn” or rugosity varies considerably in other tyrannosaurids. This is in part ontogenetic, with larger more mature specimens having larger, more rugose tuberosities. However, there are also taxonomic differences. In both Gorgosaurus and Albertosaurus , the tuberosity is positioned immediately behind the orbital margin. In Daspletosaurus sp. (FMNH PR365, MOR 590, TMP 85.62.1), the postorbital “horn” is a thickened rugose ridge that runs along the anterodorsal margin of the postorbital to the middle of the postorbital bar where it curves down and forward. The rugosity therefore looks like an open “C” that faces the orbit and encloses a smooth area in the position where the tuberosity would be in albertosaurines. This is also the case in Tyrannosaurus and Tarbosaurus . The two known specimens of Daspletosaurus torosus (NMC 9506, TMP 2001.36.1) have a pronounced convex tuberosity in this region, as apparently does OMNH 10131 ( Lehman and Carpenter 1990). The postorbitals of the immature Daspletosaurus ( Fig. 22A View Fig ) and Nanotyrannus , on the other hand, are smoothly convex.
The intertemporal ramus curves down and back from the postorbital ramus to end in a point that is clasped dorsally and ventromedially by two processes of the squamosal. Medial to the dorsolateral margin of the intertemporal ramus, the postorbital forms the smooth, gently convex floor of the superior temporal fenestra. Anteriorly, the frontal suture of the immature G. libratus (TMP 91.36.500) is nearly vertical and faces anteromedially. Behind a distinct inflection, the frontalpostorbital contact becomes more or less parallel with the midline of the skull when viewed in dorsal aspect. In this region the postorbital overlaps a wide shelf on the frontal. Posteriorly the suture turns medially where the postorbital has relatively small contacts with the parietal and laterosphenoid. In larger individuals, the anterior part of the suture becomes deeper and more rugose than the overlapping suture behind it. The suture for the frontal, parietal and laterosphenoid is essentially the same in Albertosaurus ( Fig. 8B View Fig ) and Daspletosaurus ( Fig. 22B View Fig ).
At the top of the jugal−postorbital contact in G. libratus (TMP 91.36.500), the axis of the postorbital bar has a pronounced anteroventral inflection. The orbital margin is thick dorsally, but thins ventrally. The ventral end of the postorbital ramus of the postorbital tapers rapidly and protrudes slightly into the orbit of G. libratus ( Fig. 2A View Fig , ROM 1247, TMP 86.144.1, USNM 12814). This seems to be an ontogenetic and size dependent character, and larger specimens of both G. libratus (AMNH 5336, UA 10) and A. sarcophagus (TMP 86.64.1, TMP 81.10.1) have more conspicuous suborbital processes. Suborbital processes are not found in Nanotyrannus and Alioramus, presumably because of their small size, nor in Daspletosaurus specimens of any size ( Fig. 22 View Fig , FMNH PR308, NMC 8506, TMP 84.60.1, TMP 85.62.1, 2001.36.1). However, they are present in all mature specimens of Tarbosaurus and Tyrannosaurus .
Jugal.—Tyrannosaurid jugals take part in the rim of the antorbital fenestra. In the immature Gorgosaurus ( Fig. 2B View Fig ), the jugal separates the maxilla and lacrimal by 17 mm on the right side, but only 4.5 mm on the left. The tapering anterior end of the jugal sits on the dorsal surface of the part of the ventral margin of the antorbital fossa formed by the maxilla. It overlaps the lateral surface of the maxilla here, but passes dorsally through a slot to the medial surface of the maxilla at the edge of the antorbital fenestra. Here it extends forward to contact the palatine. Posterior to this point, the medial surface of the jugal sits on the dorsolateral surface of the palatal shelf of the maxilla, and the top of the tapering end of the maxilla sits in a slot along the ventral margin of the jugal. The nature of the contacts is essentially the same in Albertosaurus ( Fig. 6 View Fig ).
Posterior to the termination of the maxilla, the ventral margin of the jugal attains its most ventral extension (the cornual process of Carr 1999) directly beneath the orbit. The cornual rugosity is anteroposteriorly shorter but dorsoventrally deeper and thicker in Daspletosaurus and Nanotyrannus than it is in larger specimens of Albertosaurus and Gorgosaurus .
The part of the orbital margin formed by the jugal ends anteriorly in a small fingerlike process within which the lacrimal is locked in place in Gorgosaurus ( Fig. 2A View Fig ) and Albertosaurus ( Fig. 6A View Fig ). Anterior to this, the jugal has a thin, plate−like anterodorsal process that fits between the outer and inner walls of the lower end of the lacrimal (AMNH 5432). Laterally this suture extends anterodorsally to the edge of the antorbital fossa.
Tyrannosaurids all have a large pneumatopore in the posteroventral corner of the antorbital fossa. The broken surfaces of TMP 82.13.3 ( A. sarcophagus ) show that pneumatization did not extend into either the postorbital or the subtemporal processes. The pneumatopore is long but slit−like in Gorgosaurus , extending most of the way from the lacrimal suture to the ventral margin of the antorbital fossa. In G. libratus (TMP 86.144.1 and UA 10), the opening is wider ventrally than dorsally, giving a teardrop shape in anterior view. When viewed laterally, the axis of the opening is inclined at an angle of about 45 degrees from the ventral margin of the skull. In contrast, the same axes are horizontal in the relatively smaller openings of Daspletosaurus ( Fig. 18A, B View Fig ), Nanotyrannus , and Tarbosaurus . As pointed out by Carr (1999), the posterolateral margin is resorbed in larger specimens to expose more of the opening in lateral view. The pneumatic opening in the posteroventral corner of the antorbital fossa is rounder in outline and lower in position in Daspletosaurus than it is in Albertosaurus and Gorgosaurus , where it is taller and slit−like. In Nanotyrannus the pneumatic opening is an almost horizontal slit.
The jugals of Albertosaurus and Gorgosaurus show significant differences from that of Daspletosaurus . The suborbital bar is both relatively and absolutely lower in Daspletosaurus (74.5 mm in TMP 85.62.1 compared with 90mm in the UA 10, which is a slightly smaller G. libratus ), but compensates for the difference in strength by being thicker (25 mm in TMP 81.24.5 compared with 16.5 mm in TMP 81.10.1, A. sarcophagus ). The ventral margin of the postorbital−jugal contact is higher in relation to the bottom of the orbit in even small specimens of Daspletosaurus than it is in either Gorgosaurus or Nanotyrannus .
The postorbital process of the jugal of the immature G. libratus (TMP 91.36.500) forms only the bottom of the orbital margin, and is excluded from 90percent of the orbit by the postorbital. Carr (1999) has shown that in larger specimens of Gorgosaurus , the anteroventral postorbital−jugal contact is higher than the ventral orbital margin. The ventral edge of the postorbital sits on a rugose, thin shelf of bone on the jugal that is 12 mm long anteroposteriorly in the immature G. libratus (TMP 91.36.500). This shelf is not evident in TMP 86.144.1 ( G. libratus ), and appears to have been something that developed ontogenetically because it is accentuated in larger specimens of Gorgosaurus and Albertosaurus . The base of the postorbital process of Albertosaurus and Gorgosaurus is shorter anteroposteriorly and the outer surface in all but the largest specimens is generally convex in longitudinal section. Juvenile Daspletosaurus ( Figs. 18A View Fig , 23 View Fig ) is similar to most albertosaurines, but larger Daspletosaurus have postorbital processes with much longer bases, each of which has a conspicuous lateral depression. In Nanotyrannus the base is broader as in Daspletosaurus , but lacks the outer depression, suggesting that the character is ontogenetically controlled as in Gorgosaurus ( Carr 1999) .
The elongate subtemporal process of the jugal divides into two processes, the longer ventral fork supporting the quadratojugal from below, and the dorsal fork forming the anteroventral margin of the lateral temporal fenestra. The quadratojugal laterally overlaps the jugal on the margins of the slit between the two forks, but to a much greater degree on the ventral fork. The anterior end of the quadratojugal suture tapers and thins anteriorly in most specimens of Gorgosaurus and Albertosaurus , whereas in all tyrannosaurines it ends in a high, double−pointed recess buttressed anteriorly by a thickening on the lateral surface of the jugal ( Fig. 23A View Fig ).
Squamosal.—Tyrannosaurid squamosals are complex but conservative. The lateral surface of the intertemporal ramus of the squamosal is lightly sculptured above the long, slit−like articulation for the postorbital ( Fig. 2A View Fig ). The two branches of the intertemporal ramus both extend to the anterior margin of the lateral temporal fenestra, even though the lower one is hidden laterally by the postorbital. Isolated G. libratus specimens (TMP 86.144.1, TMP 92.36.82) show that the squamosal medially overlaps the posterior half of the intertemporal ramus of the postorbital. The squamosal of Gorgosaurus has a long tapering process that runs forward, curving somewhat downward, along the top of the anterodorsal ala of the quadratojugal. Together they almost dissect the lateral temporal fenestra. This quadratojugal process is relatively thick along its dorsal margin, and is almost completely exposed in lateral view. It is not extensively overlapped laterally as it is in Alioramus ( Kurzanov 1976) , Daspletosaurus sp. ( Fig. 25 View Fig , TMP 85.62.1) and Tarbosaurus bataar (GIN 107/2). Surrounded by most of the processes of the squamosal, the ventral surface is deeply concave. The juvenile specimen of Daspletosaurus also has a deep concavity with relatively poorly defined margins ( Fig. 25D View Fig ). In mature specimens of Gorgosaurus , Albertosaurus , and all tyrannosaurines, the concavity has a well−defined margin and extends deeply into the interior, strongly suggesting it housed an air sac.
Quadratojugal.—The quadratojugal ( Fig. 2A View Fig ) expands dorsally into a wing like process along its dorsal contact with the squamosal. At the back of the dorsal margin, the quadratojugal pulls away from the squamosal and contacts a ridge of the quadrate anteroventral to the quadrate cotylus. The quadratojugal thickens slightly along its sutural contact, which extends ventrally into a slot. The quadrate passes through the slot, and the quadratojugal wraps around onto the posterolateral surface of the quadrate, overlapping the quadrate extensively above the quadrate fenestra. The quadratojugal forms the lateral margin of the large quadrate fenestra, and contacts the quadrate again ventral to the opening. Medially, the quadrate suture of G. libratus (TMP 86.144.1) and Albertosaurus ( Fig. 9B View Fig ) is concave and rugose, matching in contours the extensive sutural surface on the quadrate. The anterior process of the quadratojugal extensively overlaps the lateral surface of the lower of the two quadratojugal processes of the jugal.
The quadratojugals of Albertosaurus and Gorgosaurus are quite different from that of Daspletosaurus ( Fig. 24 View Fig , NMC 8506, TMP 85.62.1) and other tyrannosaurines, in which the dorsal contact with the squamosal is anteroposteriorly much more limited. The anterior and posterior margins of the ascending part of the quadratojugal are relatively straight in lateral view as they diverge from each other dorsally in tyrannosaurines.
Quadrate.—Tyrannosaurid quadrates ( Figs. 10 View Fig , 28 View Fig ) are relatively short in comparison with most other theropods. The quadrate has a large pneumatopore ( Fig. 28C View Fig ) on the ventral margin of the base of the pterygoid ala. The medial surface of the pterygoid ala has a well−defined concavity ( Figs. 10B View Fig , 28B View Fig ) anterodorsal to the pneumatopore that may mark the position of an air sac. It is bound anterodorsally, anteriorly and anteroventrally by the curved, squamose suture for the quadrate ala of the pterygoid. The quadrate cotylus ( Fig. 28D View Fig ) contacted only the squamosal. However, a small, flattened area on the medioposterior surface of the quadrate ( Fig. 28B View Fig ) just below the cotylar surface abutted against the opisthotic−exoccipital.
Occiput.—On the occipital surface of the immature G. libratus (TMP 91.36.500), the foramen magnum is less than half the diameter of the nearly circular occipital condyle. The sutures on the occiput are visible, but are better seen on TMP 86.144.1 ( G. libratus ) and 94.143.1 ( Daspletosaurus sp.) where the bones have separated.
Supraoccipital.—The supraoccipital forms a narrow part of the dorsal margin of the foramen magnum in G. libratus (TMP 86.144.1, MOR 557, TMP 94.12.602), A. sarcophagus (TMP 81.10.1), Daspletosaurus ( Fig. 26B View Fig ) and Tyrannosaurus ( Brochu 2002) . The assumption that the exoccipitals connect across the top of the foramen magnum in tyrannosaurids ( Holtz 2001) is probably incorrect for all genera. A strong midline ridge rises above the foramen magnum, and the external occipital vein exits through a depression on each side. Tyrannosaurids have a knob−like process on each side of the midline at the top of the supraoccipital. In large specimens of G. libratus (TMP 94.12.602), these protrude more than 3 cm from the contact with the parietal. Numerous tyrannosaurid specimens have broken supraoccipitals that reveal internal sinuses. Most of these have relatively thick walls, but in TMP 94.143.1 ( Daspletosaurus sp.) two millimeter thick walls enclose an internal chamber that seems to be confluent with the sinuses in the paroccipital processes. The pneumaticity of a much larger Tyrannosaurus braincase ( Brochu 2002) with 3 mm thick walls is even more impressive. The chamber is partially separated by a vertical midline ridge on the anterior wall. The pneumaticity of the supraoccipital is reminiscent of Troodon ( Currie and Zhao, 1993 B). Basioccipital.—The basioccipital narrowly separates the exoccipitals on top of the occipital condyle. The condyle in even large specimens of Gorgosaurus is oriented mostly posteriorly (TMP 94.12.602), but faces somewhat downward as well. Just outside of the articular surface, the neck of the G. libratus condyle is pierced ventrally by a single foramen, which is on the left side of TMP 86.144.1 and on the right side in TMP 91.36.500. This lack of consistency suggests that the foramen was a pneumatopore. A midline ridge supporting the condyle divides ventrally, and each resulting ridge ends at one of the basal tubera. There is a large depression lateral to each one of these ridges, which housed an air sac. Pneumatopores penetrate into the interior of the basioccipital in the dorsal region of this depression in NMC 5600 ( A. sarcophagus ), ROM 1247 ( G. libratus ), and TMP 81.10.1 ( A. sarcophagus ). This depression extends over a low ridge onto the posterior surface of the exoccipital, where one or more pneumatopores invade the interior of that bone as well. Ventrally, the basioccipital forms most of the basal tubera, which are supported anteriorly by the basisphenoid. Anterolaterally, the metotic strut of the exoccipital−opisthotic almost reaches the ventral surface of the basal tuber. The width across the basal tubera is greater than width of the occipital condyle (25% greater in TMP 91.36.500, G. libratus ). However, this is still relatively narrow in comparison with other tyrannosaurid genera. The height of the basioccipital below the condyle on the midline does not increase as much as the distance to the end of the basioccipital tuber with increasing age in Albertosaurus and Gorgosaurus . The ratio of basal tuber height to midline height is 1.4 in TMP 86.144.1 ( G. libratus ), 1.9 in TMP 91.36.500 ( G. libratus ), and 2.5 in TMP 81.10.1 ( A. sarcophagus ).
Exoccipital−opisthotic.—Ventrally, the metotic strut is a triangular plate of bone that contacts the basioccipital medially, but is overlapped slightly by it distally. Anterolaterally, the basisphenoid overlaps about a quarter of the metotic strut. Posteromedially, there is a shallow depression with a pneumatopore that seems to penetrate into the base of the exoccipital contribution to the occipital condyle, and another that passes through the suture with the basisphenoid into the interior of that bone. The paroccipital process is oriented laterally, posteriorly and only slightly downward from the foramen magnum. Two foramina ( Figs. 26 View Fig , 27 View Fig ) pierce the anterolateral surface of the unit behind the fenestra ovalis. As they are not connected with any of the other foramina, it appears they might be pneumatopores that took air to the interior of the paroccipital process, making them equivalent to the posterior tympanic sinuses of troodontids and other theropods. This is particularly evident in TMP 94.143.1 ( Daspletosaurus sp.), where the left paroccipital has been broken to expose the hollow interior ( Fig. 26 View Fig ) that connects via tubes to the anterior tympanic recess and the hollow interior of the supraoccipital as in Troodon ( Currie and Zhao 1993b) . The exoccipital−opisthotic unit forms the dorsal, posterior and ventral edges of the fenestra ovalis, the border being completed by the prootic and the basisphenoid. Above the fenestra ovalis, the bone thickens in TMP 86.144.1 ( G. libratus ) where it encloses the back regions of the inner ear. Posterolateral to this, the anterolateral surface of the exoccipital−opisthotic is broadly overlapped by the prootic. Dorsally, the exoccipital−opisthotic has an anteroventrally long suture with the supraoccipital−epiotic, which roofs over the inner ear. Posterolateral to this, the dorsal edge of the unit thins out along its contacts with the parietal and squamosal. Where the exoccipital−opisthotic forms the inner wall at the back of the braincase, there is a foramen for the ductus endolymphaticus behind the metotic fissure.
Basisphenoid−parasphenoid.—The cultriform process extends anteriorly just beyond the middle of the orbit, where it is a thin but solid sheet of bone. The well developed basipterygoid process inserts into the socket−like pterygoid articulation. A web of bone connects the bases of the basipterygoid processes. As in other specimens of G. libratus and A. sarcophagus (NMC 5600, ROM 1247, TMP 81.10.1, TMP 85.98.1, 86.64.1), the ventrally facing basipterygoid recess of TMP 91.36.500 ( G. libratus ) is subdivided into a large medial, anterior depression, and a pair of posterolateral pits. This is different than the broader, shallower basisphenoid recesses of Alioramus remotus (PIN 3141), Daspletosaurus sp. (FMNH PR308, NMC 8506, Fig. 26C View Fig ), Nanotyrannus , Tarbosaurus bataar (PIN 553−3), and Tyrannosaurus , in all of which the basisphenoidal recesses are also oriented more posteroventrally. The ala basisphenoidalis ( Chure and Madsen 1998) enclosed the anterior part of the air sac associated with the middle ear, and overhangs the foramen for the internal carotid.
Prootic.—The prootic in tyrannosaurids ( Fig. 6B View Fig , G. libratus TMP 86.144.1) forms the posterior border for the opening of the trigeminal, the anterior margin of the fenestra ovalis, and completely encloses the foramina for the facial and auditory nerves. Posterolaterally the prootic extensively overlaps the anterolateral face of the opisthotic.
Laterosphenoid.—As in other theropods, the tyrannosaurid laterosphenoid forms the anterior margin of the trigeminal foramen, and there was a separate branch for the ophthalmic branch of the trigeminal. Dorsally, the postorbital process of the laterosphenoid contacted the frontoparietal suture close to where all three bones meet the postorbital ( Fig. 6B View Fig ). There is a shallow depression on the ventrolateral surface of the base of the postorbital process marking the contact with the epipterygoid.
Vomer.—Tyrannosaurid vomers, which are fused for most of their lengths, are distinctive in that each expands anteriorly into a relatively large, diamond−shaped plate that loosely contacts the lower surface of the palatal shelf of the maxilla. Posteriorly, the vomer splits into two vertical plates that are narrowly separated along the midline and contact the palatines and pterygoids posterolaterally in G. libratus (TMP 2000.12.11), A. sarcophagus (TMP 98.63.87), Daspletosaurus ( Russell 1970) , Tarbosaurus bataar (GIN 100/777, GIN 107/2), and Tyrannosaurus ( Osborn 1912, Molnar 1991, Brochu 2002).
Palatine.—The ventral edge of the palatine is sutured to the medial edge of the palatal shelf of the maxilla for most of its length. Behind the last alveolus, this sutural surface shifts dorsally onto the jugal, where it becomes tall enough to contact the medial surface of the lacrimal (TMP 91.36.500 [ G. libratus ], CM 7541 [ N. lancensis ]). There is one ( Fig. 11A View Fig , CM 7541) or more ( Fig. 29 View Fig ) pneumatic openings above the maxillary suture on the lateral surface. The fenestra shows a general trend towards increasing relative size in larger individuals, and taxonomically seems to be relatively larger in Albertosaurus , Gorgosaurus and Nanotyrannus than it is in Daspletosaurus . The pneumatic space extends dorsomedially into the inflated base of the vomerine process. Anterior to the pneumatic fenestra, the lateral surface of the tapering maxillary process of the palatine has a deep concavity above the maxillary suture that also housed a pneumatic sac. The vomerine process expands dorsally ( Figs. 2A View Fig , 11A View Fig ) into a large, thin, almost vertical sheet of bone that covers the lateral surface of the vomerine process of the pterygoid. The palatines of Gorgosaurus and Albertosaurus , and of all tyrannosaurine juveniles are relatively flat medial to the pneumatopore. However, the base of the palatine becomes greatly inflated in large specimens of Daspletosaurus sp. (TMP 85.62.1), Tarbosaurus and T. rex (MOR 008, 555).
Ectopterygoid.—Tyrannosaurid ectopterygoids ( Figs. 2B View Fig , 12 View Fig , 32 View Fig ) are conservative in their features. The hook−like jugal process expands distally for its contact with the medial surface for the jugal, and just touches the maxilla anteroventrally. Dorsally the ectopterygoid develops into a broad plate that has an extensive squamose suture with the lateral surface of the pterygoid. The posterodorsal corner of this process is thickened on the ectopterygoid to form a process that plugs into a socket in the pterygoid. The more ventromedial, hooked process of the ectopterygoid protrudes below the ventral margin of the skull. The process is thick anterolaterally, and forms the margin of a prominent ventromedial pneumatopore, which is slit−like in Albertosaurus and Gorgosaurus . Posterior to the pneumatopore, the ectopterygoid is a thin plate of bone that is covered ventromedially by the thin, ventral process of the pterygoid. The distal end of the pterygoid wraps around onto the posterolateral face of the ventral extremity of the ectopterygoid. The ectopterygoids of tyrannosaurine juveniles are difficult to distinguish from those of albertosaurines. However, as a tyrannosaurine matured and increased in size, the base of the ectopterygoid became inflated.
Pterygoid.—The pterygoids are both well exposed in the immature Gorgosaurus ( Fig. 2B View Fig ), and extend for slightly less than half the length of the skull. As pointed out by Russell (1970), tyrannosaurid pterygoids do not contact each other on the midline, and are separated anteriorly by the paired vertical plates of the paired vomers. The pterygoids of Gorgosaurus and Albertosaurus are similar in most respects to those of Daspletosaurus , Tarbosaurus ( Maleev 1974) , Nanotyrannus , and T. rex (BHI 3033, Osborn 1912; Molnar 1991), except that they are somewhat more elongate between the basipterygoid articulation and the ectopterygoid process, and the anterior ramus is not as broad as in tyrannosaurines.
The anterodorsal ramus matches the shape of the dorsomedial process of the palatine, which overlaps the pterygoid in an extensive squamose suture. The pterygoid narrows posteriorly, separates from the palatine and forms the posterior margin for an accessory opening between the two bones. In Albertosaurus ( Fig. 13 View Fig ), the accessory fenestra is readily visible in ventral view, whereas in TMP 91.36.500 ( G. libratus ) it is half the size. It is also a relatively large, anteroposteriorly elongate opening in the juvenile specimen of Daspletosaurus ( Fig. 29B View Fig ), and apparently in Nanotyrannus . It is supposedly closed in adult specimens of Tyrannosaurus ( Osborn 1912, Molnar 1991, Carr 1999), as a result of the anterior process of the pterygoid becoming relatively wider and because of medial expansion of the palatine. However, it never closes completely, and in BHI 3033 is still a sizeable opening. Because of overlapping bones, it may not be visible in ventral view. Posteroventral to this fenestra, there is an anteroventral projection that forms a squamose suture along the medial surface of the palatine that ends anteriorly at a thickened palatine ridge. The palatine extends far back along the lateral surface of the pterygoid, but does not contact the ectopterygoid (the contact in NMC 5600 is probably postmortem damage). The ventral border of the pterygoid is thickened below the palatine contact. There is a long fingerlike process that extends ventrally, laterally and slightly posteriorly along the posterior edge of the ventral ramus of the ectopterygoid. Because of the great posterior width of the skull, the process is relatively longer in the smaller Nanotyrannus skull than it is in the immature G. libratus (TMP 91.36.500). Near the base of the process, the pterygoid covers the ventral surface of the back of the ectopterygoid, but distally it twists around the posterior edge of the ectopterygoid to terminate on the dorsal surface. Dorsolaterally, the ectopterygoid contact extends from the base of the ventral process to plug into an anterolaterally facing socket at the base of the quadrate ala of the pterygoid. The quadrate ramus is a thin, curved, quadrilateral sheet of bone that is reinforced and strengthened by heavy ridges along the medial surface of the anterodorsal margin, and the lateral surface of the anteroventral edge. The more dorsal of these two ridges ends in a suture for the epipterygoid, which also covers part of the lateral surface of the quadrate ramus of the pterygoid. Almost half of the somewhat convex, lateral surface of the quadrate ala is covered by a squamose sutural contact with the quadrate.
Because the skulls of tyrannosaurines are wider behind the orbit, the quadrate process of the pterygoid is longer and more laterally oriented in tyrannosaurines.
Epipterygoid.—Epipterygoids are well preserved in two of the juvenile Gorgosaurus . Both are in position in TMP 91.36.500, whereas they are disarticulated in TMP 86.144.1 ( G. libratus ). Lambe (1904) described the upper part of the epipterygoid of Albertosaurus , which is essentially the same as an incomplete epipterygoid in Daspletosaurus ( Fig. 31 View Fig ). One large Daspletosaurus (TMP 85.62.1) has an epipterygoid that is comparable with those reported for Tyrannosaurus ( Osborn 1912; Molnar 1991). The epipterygoid tapers dorsally, and forks ventrally into an anterior process continuous with the anterior ridge of the quadrate process of the pterygoid, and a posterior process that extends posteriorly along the dorsal edge of the pterygoid. Both processes overlap the lateral surface of the quadrate process, although the larger, more posterior fork has a more extensive area of contact. The outer surface of the epipterygoid has a shallow concavity near its base. Dorsally, there is an anteromedially oriented facet for contact with the laterosphenoid.
Stapes.—In a large specimen of G. libratus (TMP 94.12.602), the preserved shaft of the stapes is 90mm long and has a maximum shaft diameter of 2.5 mm. The stapes of Tyrannosaurus ( Brochu 2002) is also about 2 mm in diameter.
Lower jaw
Mandible.—In the immature G. libratus (TMP 91.36.500), the posteriorly projecting intramandibular process of the dentary fits into a depression on the lateral surface of an anterodorsal, tongue−like process of the surangular. The external mandibular fenestra has irregular margins but is almost oval in shape. Ventrally, the anterior end of the angular slides between the dentary and splenial, separating them for a short distance where the posterior tips of the two anterior bones fit into grooves on either side of the angular. Although there appears to have been considerable sliding movement possible in the ventral part of the intramandibular joint, the complex dorsal joint between the dentary and the surangular would have only permitted the two bones a range of about 1.5 cm before they interlocked to form a pivot for any further motion. The upper part of the intramandibular joint is also stabilized by the coronoid−supradentary ossification that spans the joint. The coronoid−supradentary seems to have been capable of some limited sliding motion in a groove on the medial surface of the surangular, and is so thin that it may have been capable of some bending. The intramandibular joint of Gorgosaurus is typical of tyrannosaurids, although the flexibility was reduced in Tarbosaurus ( Hurum and Currie 2000) and possibly the other large tyrannosaurines. Differences in the intramandibular joints may reflect differences in feeding behavior ( Hurum and Currie 2000, Hurum and Sabath 2003).
Dentary.—The lateral surface of the dentary is pierced by a dorsal row of mental foramina for innervation of the skin and lips by the inferior alveolar nerve. A row of smaller, less prominent foramina is found near the ventral margin of the lateral surface of the dentary. The interdentary symphysis is poorly defined in most tyrannosaurid jaws, and allowed some intermandibular movement. Unlike the juvenile Daspletosaurus sp. ( Fig. 33 View Fig ), one large specimen of D. torosus (NMC 8506) has a complex, rugose symphysis, which suggests that movement would not have been possible between the mandibles. This specimen also shows some coossification of the premaxillae, which is not evident in another D. torosus specimen (TMP 2001.36.1) that was just as large an individual. The heavy ossification of the symphysis and premaxillae of NMC 8506 may indicate a health problem.
All well−preserved tyrannosaurid dentaries have three posterior processes—the intramandibular process, a process that contacts the surangular above the external mandibular fenestra, and a process that articulates with the angular below the external mandibular fenestra ( Fig. 2A View Fig ). Medially, the dental shelf is thick, and ends posteromedially in a somewhat downturned process wedged between the coronoid−supradentary laterally and the splenial medially ( Fig. 33B View Fig , TMP 95.5.1, G. libratus ). In G. libratus (TMP 91.36.500), the most anterior dentary teeth are tiny compared to the others. The interdental plates are separate from each other in all tyrannosaurids ( Fig. 33B View Fig ), but are covered medially by the coronoid−supradentary.
TMP 94.12.155 is a pair of lower jaws from a small G. libratus ( Currie and Dong 2001) , with an estimated skull length of 364 mm, which suggests that the whole animal was only about 3 m long. The dentaries are elongate (height/ length ratio of 0.10) compared to the G. libratus TMP 91.36.500 (ratio is 0.13) and larger specimens (for example it is 0.17 in UA 10, G. libratus ). Mandibular proportions are ontogenetically controlled in tyrannosaurids, and become relatively deeper in larger individuals. The lowest height of the dentary in the immature G. libratus TMP 91.36.500 is 21% the height of the length of the dentary tooth row, and 24% higher than the tallest dentary tooth. The minimum height of the similar sized Nanotyrannus is also 21% of the dentary tooth row. In the smallest known G. libratus (TMP 94.12.155), the height of the dentary is only 14% of the dentary tooth row length, and the highest dentary tooth is actually taller than the jaw beneath it. In large specimens of Albertosaurus ( Table 4 View Table4 ), the height of the jaw is about 25% the length of the tooth row, and the longest tooth is half the height of the jaw. Statistically, comparison of tooth row length versus jaw height in 27 tyrannosaur jaws ( Table 4 View Table4 ) shows that the depth of the jaw increases with strong positive allometry (k= 1.43, r 2 = 0.93). The relationship between jaw height and tooth row length ( Fig. 17 View Fig ) is consistent for both ontogenetic and interspecific size changes, demonstrating that relative jaw proportions cannot be used for distinguishing taxa.
Splenial.—In medial view, the posterior margin of the splenial is deeply notched where it forms the anterior margin of the internal mandibular fenestra. Posteriorly, the tapered end of the splenial sits in a triangular depression on the medial surface of the angular. The anterior mylohyoid fenestra ( Fig. 34 View Fig ) is almost completely surrounded by the splenial, although there is a small gap in the ventral border. Compared with most theropods, it is relatively large in tyrannosaurids. The ventral margin of the splenial is thick to support an elongate ventrolaterally oriented contact with the dentary (TMP 86.144.1, G. libratus ). Dorsal to this ridge is a groove ( Fig. 34B View Fig ) for the sliding contact with the angular. Ventral to the anterior mylohyoid fenestra, a second ventrally oriented butt joint for the dentary is positioned along the ventral margin. Anterior to the front margin of the Meckelian canal, the tapered end of the splenial medially overlaps the dentary. The dorsal butt joints with the dentary are elongate but poorly defined in the small specimens of Gorgosaurus but can be an elaborate series of anteroventrally oriented ridges and grooves in large tyrannosaurids (NMC 11594, G. libratus ).
Coronoid−supradentary.—Forming the dorsoanterior margin of the adductor fossa, the coronoid has a concave posterior margin. Posterodorsally, it has a thin, elongate butt joint with the surangular. However, more anteriorly, the coronoid twists into a well−defined pocket on the medial surface of the anterodorsal process of the surangular ( Figs. 14B View Fig , 36B View Fig ). The ventral margin of the coronoid is overlapped by the posterodorsal edge of the prearticular. The coronoid extends anteriorly as the supradentary. There is no indication of a suture or line of fusion in tyrannosaurids ( Fig. 35 View Fig ) or other theropods. The supradentary covers up the interdental plates anteriorly to at least the level of the fifth dentary tooth in the immature G. libratus (TMP 91.36.500), although most likely extends to the second dentary tooth as it does in NMC 5600. It is partially overlapped by the splenial behind the level of the last two dentary teeth.
Surangular.—The anterior surangular foramen opens anteriorly into a groove that is continuous anteriorly with the depression that receives the intramandibular process of the dentary. The large size of the posterior surangular fenestra is synapomorphic for tyrannosaurids ( Holtz 2001). The longitudinal ridge (surangular shelf) above it becomes very wide in large tyrannosaurids, and its dorsal surface has a longitudinal concavity for the powerful M. adductor mandibulae externus. The great width of the shelf is similar to the condition seen in some other groups of large theropods, including abelisaurids and carcharodontosaurids. There is an extensive squamose suture where the angular overlaps the ventral margin of the surangular behind the external mandibular fenestra ( Figs. 2 View Fig , 14 View Fig , 36 View Fig ). Behind the posterior end of the angular, the surangular forms the ventrolateral margin of the jaw. Here, it overlaps the articular, and the lateral edge of the prearticular. Posterodorsal to the posterior surangular fenestra on the medial surface there is a hook−like process ( Figs. 4 View Fig , 36B, 36C View Fig ) that contacts the lateral surface of the prearticular and forms the posterior margin of the adductor fossa. Most of the medial surface of the surangular’s contribution to the short retroarticular process is in contact with the articular. Dorsally the surangular forms half of the lateral depression of the glenoid ( Fig. 4 View Fig ) that articulates with the lateral condyle of the quadrate.
Angular.—Tyrannosaurid angulars ( Figs. 2 View Fig , 15 View Fig ) are shallowly curved plates of bone that taper and thicken antero−ventrally to form the posterior and ventral margins of the external mandibular fenestra. The posterior margin of the depressed articular facet for the dentary reaches the level of the posterior border of the external mandibular fenestra, but the medial depression for the splenial ends farther forward. Almost half of the length of the bone is covered laterally by the dentary. The ventral margin is strengthened by a ridge beneath the external mandibular fenestra, where the angular forms the ventral margin of the mandible. Posteriorly, the ventral edge rests in a slot on the lateral margin of the prearticular, and the dorsal margin overlaps the lateral surface of the surangular. The bone ends behind the posterior margin of the posterior surangular fenestra in Gorgosaurus ( Fig. 2A View Fig ), behind ( Fig. 36A View Fig ) or beneath the fenestra in Daspletosaurus ( Russell 1970) , and behind it in Nanotyrannus .
Prearticular.—The elongate tyrannosaurid prearticular expands into thin, vertical plates of bone anteriorly and posteriorly ( Fig. 16 View Fig ), but is relatively thick medially where it forms the floor of the adductor fossa and joints the angular to form the ventral margin of the jaw. The elongate but curved groove for the angular on the ventrolateral surface of the prearticular is bound dorsally by a longitudinal ridge ( Fig. 16A View Fig , 86.144.1, G. libratus ). Posteriorly, the prearticular contacts the hook−like process of the surangular behind the adductor fossa, and extensively overlaps the ventromedial surface of the articular.
Articular.—The medial glenoid and most of the interglenoid ridge for the articulation with the quadrate are found on the articular ( Fig. 4 View Fig ). The retroarticular process is short in all tyrannosaurids, and is directed posteromedially ( Fig. 2 View Fig ). The opening for the chorda tympani and posterior condylar artery ( Fig. 4 View Fig ) is large, and presumably carried a diverticulum of the tympanic air sac into the hollow core of the articular ( Molnar 1991). The anterior opening for the chorda tympani leaves the articular through the suture underneath the prearticular.
Teeth.—The number of teeth in tyrannosaurid jaws is consistently four in the premaxilla, but tends to vary in the maxilla and dentary. The variability in part is accounted for by our inability to clearly see the alveoli in some specimens. The small, anterior−most maxillary tooth is sometimes not preserved because this part of the maxilla is easily damaged, or it can be counted as a premaxillary tooth if the suture is not clean enough to be visible. This happens easily because of its small size, incisiform shape and position at the premaxillary−maxillary suture.
Regardless of these problems, maxillary and dentary tooth counts are still variable within any taxon. In fact they can even vary from one side of the mouth to the other. For example, TMP 94.12.602 ( G. libratus ) has 14 maxillary teeth on the right side and 15 on the left, TMP 91.36.500 ( G. libratus ) and NMC 11594 ( Daspletosaurus sp.) have 16 dentary teeth on one side and 17 on the other, GIN 107/1 ( Tarbosaurus bataar ) has 12 and 13 alveoli in the right and left maxillae, and SDSM 12047 ( T. rex ) has 11 left and 12 right maxillary teeth ( Molnar 1991). Maxillary tooth counts vary between taxa, and amongst different individuals within a genus. Most specimens of G. libratus have 13 or 14 maxillary teeth, but some have 15 (NMC 2270, TMP 83.36.100, 94.12.602). The same range exists in Albertosaurus . In Daspletosaurus sp., TMP 94.143.1 ( Fig. 18A View Fig ) has only 13 maxillary alveoli whereas all other specimens from Alberta have 15 or 16, and those from Montana have as many as 17 (MOR 590). Anatomical, geographic and stratigraphic differences, however, suggest that there are as many as three species of Daspletosaurus . All Tarbosaurus bataar specimens have either 12 or 13 maxillary teeth (GIN 107/1 has 12 in one maxilla and 13 in the other), and T. rex has 11 or 12 (SDSM 12047 has both counts). Dentary teeth show the same kind of variability, although there are usually one or two more alveoli in the dentary than there are in the maxilla of the same individual.
There are several synapomorphies in the premaxillary teeth of tyrannosaurids ( Holtz 2001). First, they are always significantly smaller than the largest maxillary or dentary teeth ( Figs. 2A View Fig , 18A, 18C View Fig ). More significant for the recovery of identifiable isolated teeth, however, is the fact that they are D−shaped in cross−section (Currie et. al 1990). In tyrannosaurid premaxillary teeth, both carina are on the posterior surface of the tooth, and in most cases the maximum mediolateral width of the tooth is greater than the distance between the two carina. All premaxillary teeth in Gorgosaurus are serrated, in contrast with “ Aublysodon ”. The holotype of “ Stygivenator molnari ” (LACM 28471), an immature specimen of T. rex , lacks serrations on the only premaxillary tooth preserved ( Molnar 1978). The juvenile specimen of Daspletosaurus has no serrations on either carina, although its first incisiform maxillary tooth has denticles.
Of the fourteen maxillary teeth in the immature G. libratus (TMP 91.36.500), the first is incisiform and is as tall as the longest premaxillary tooth. The second maxillary tooth is transitional in size and shape, although it is no longer D−shaped in cross−section. By the third maxillary tooth, the anterior carina is positioned on the midline distally, but is closer to the lingual side of the tooth proximally. The posterior carina runs along the posterolabial edge of the tooth. The longest maxillary teeth are in the fourth to eighth alveoli. The last maxillary tooth is only a third of that height.
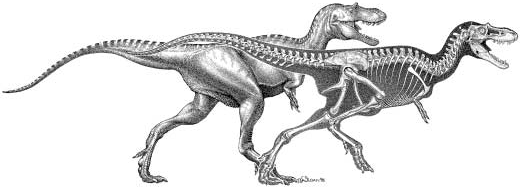
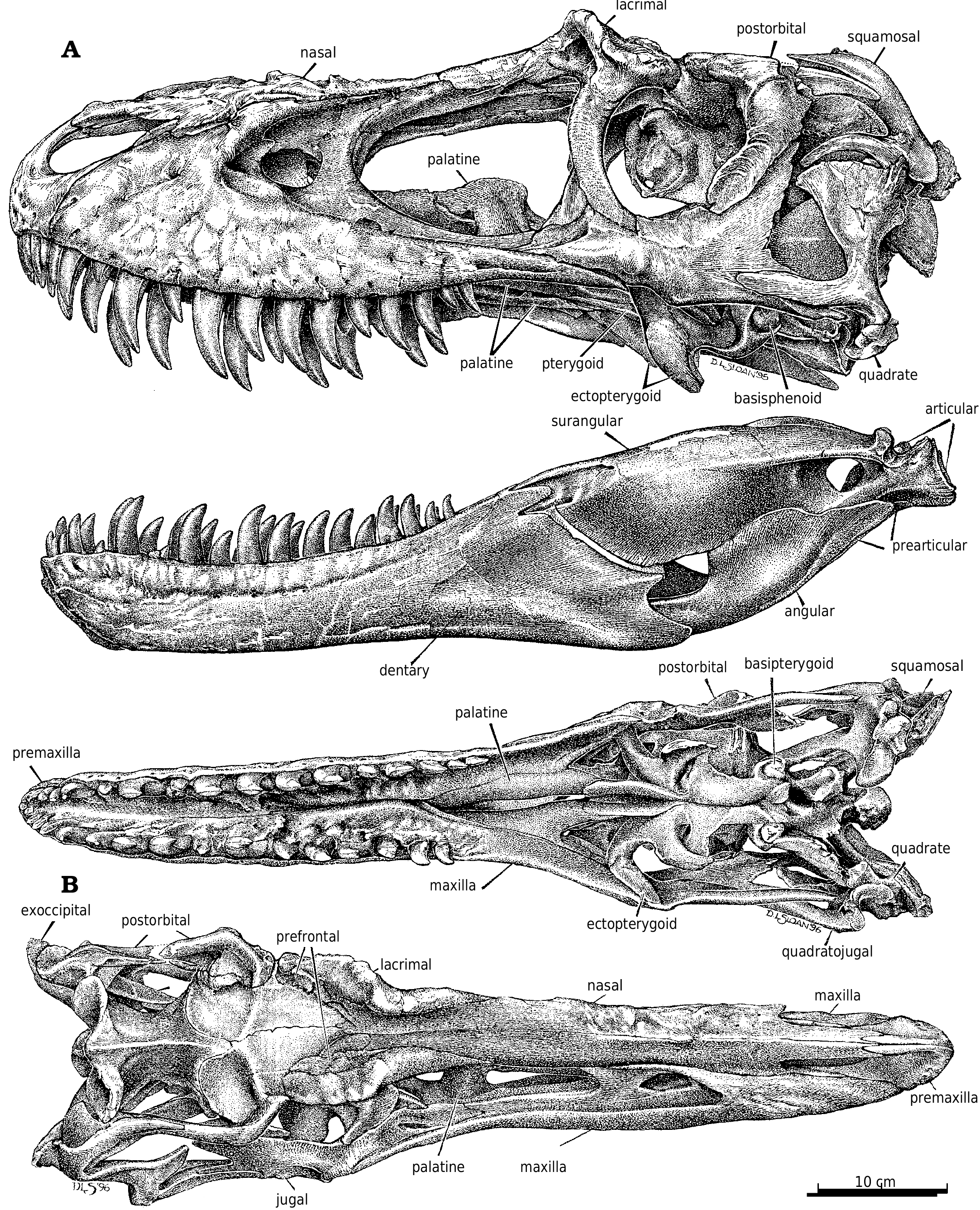
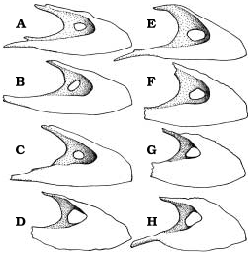
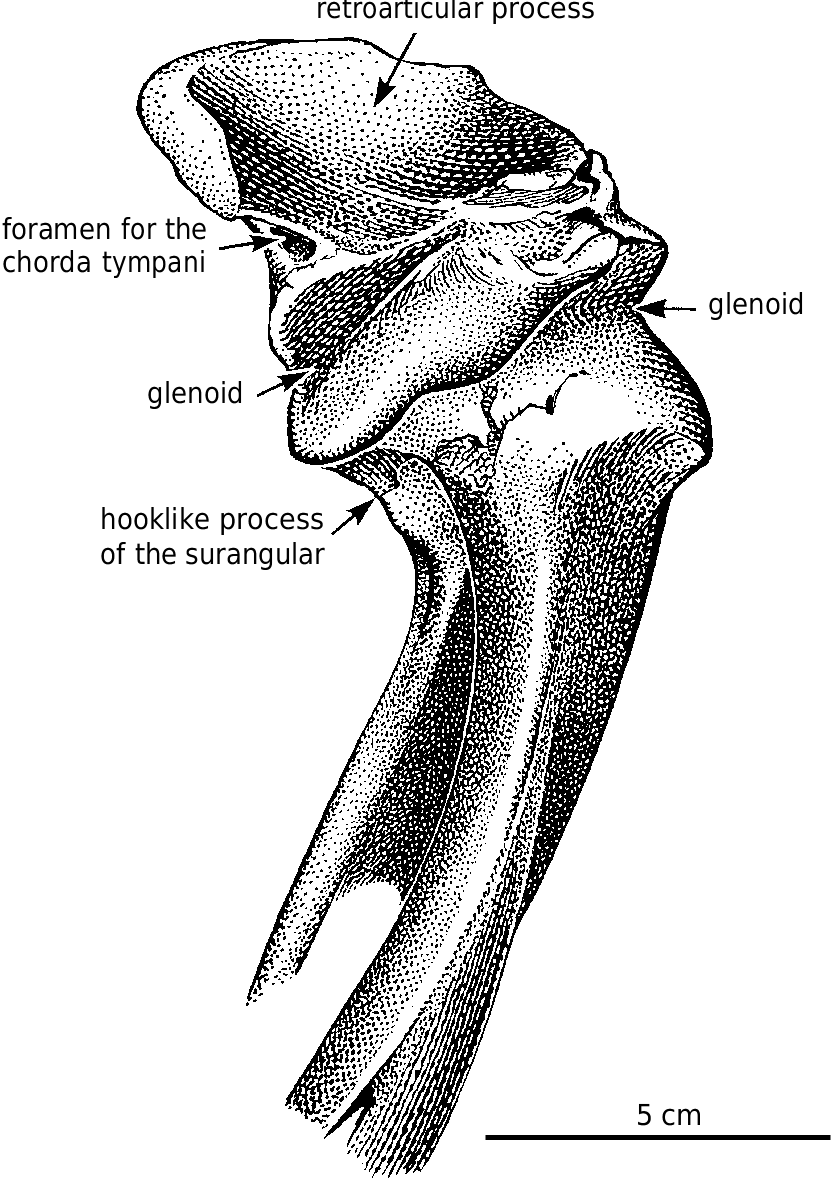
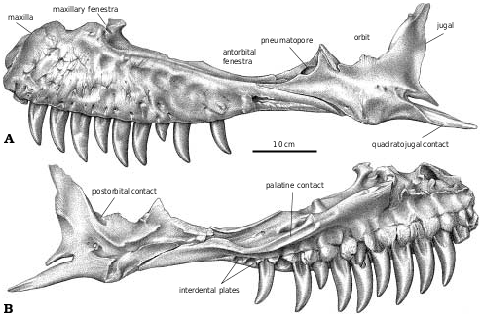
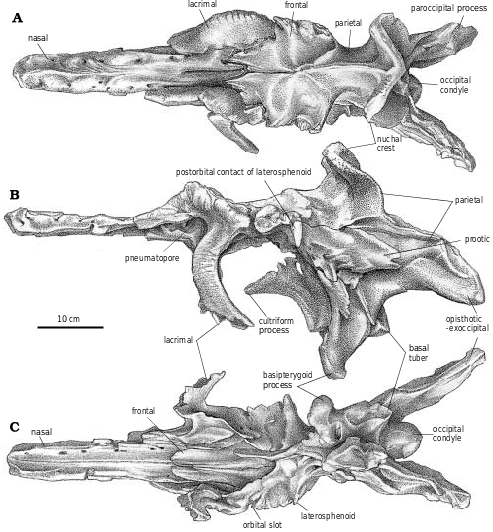
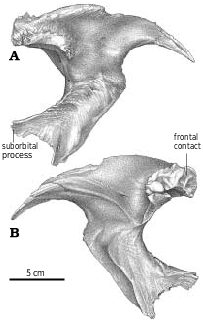
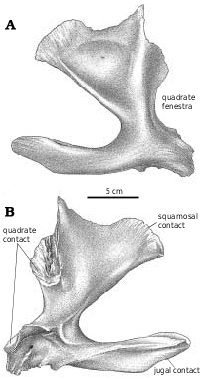
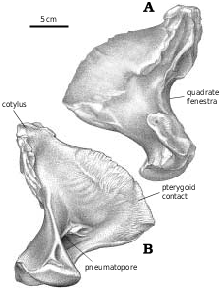
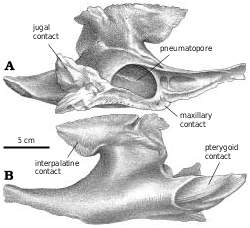
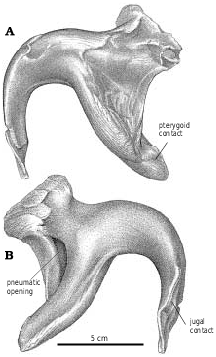
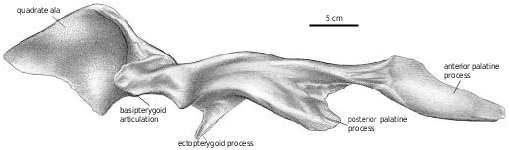
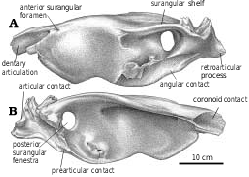
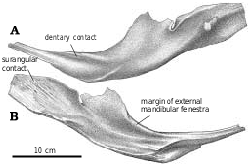
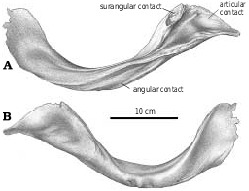

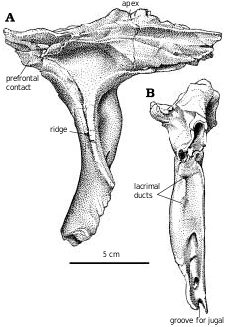
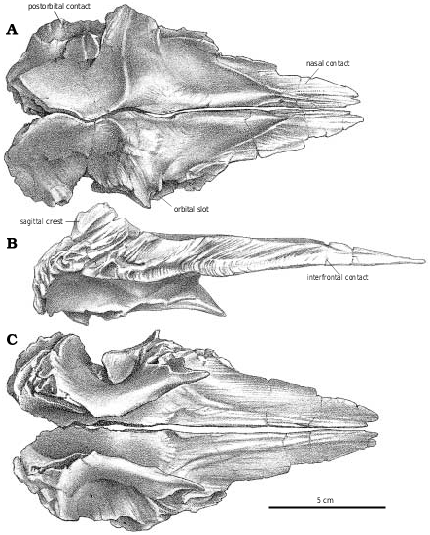

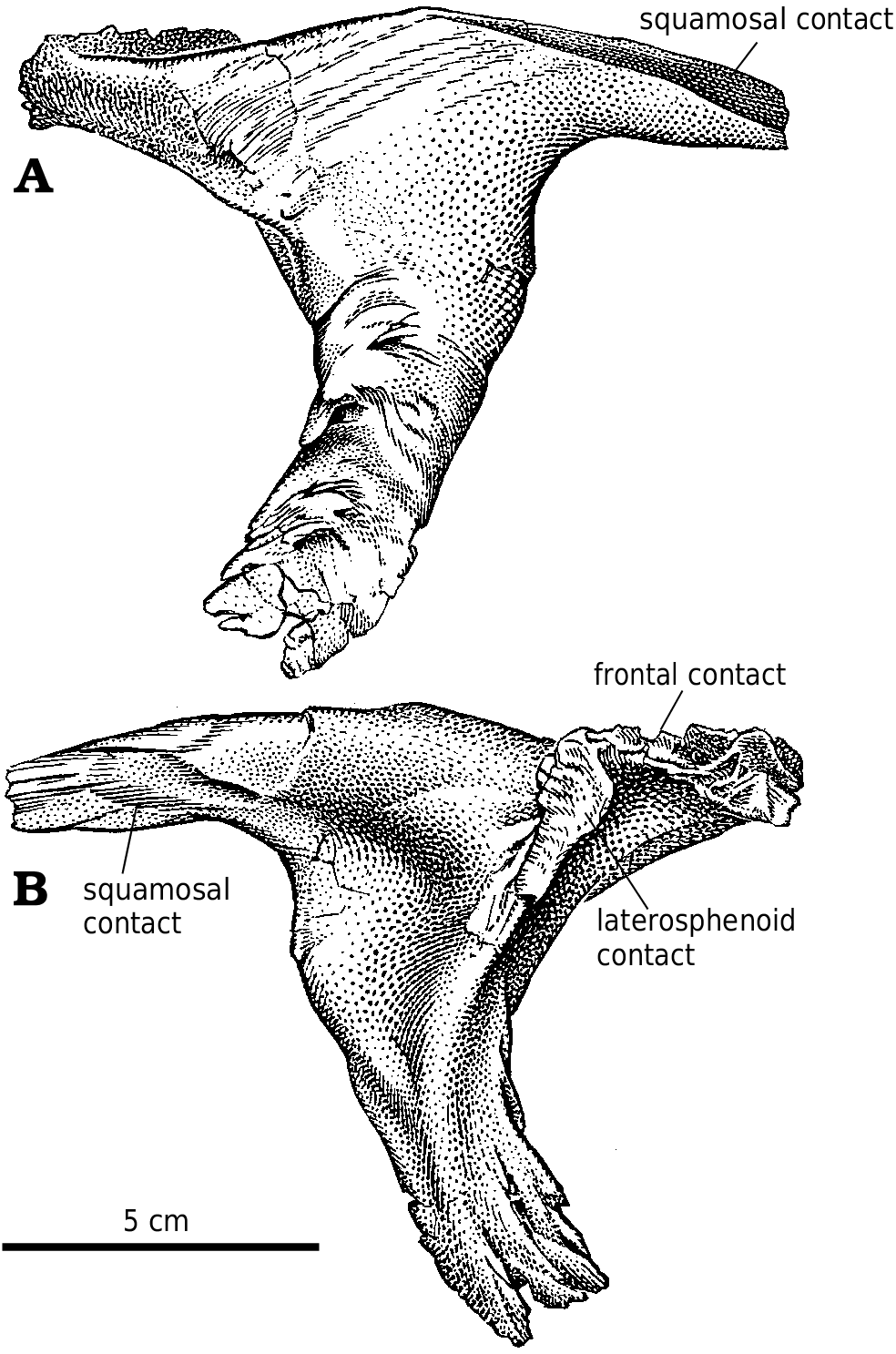
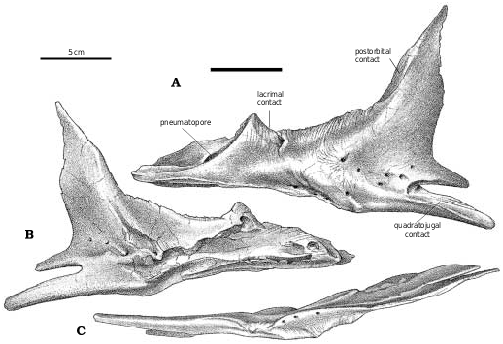
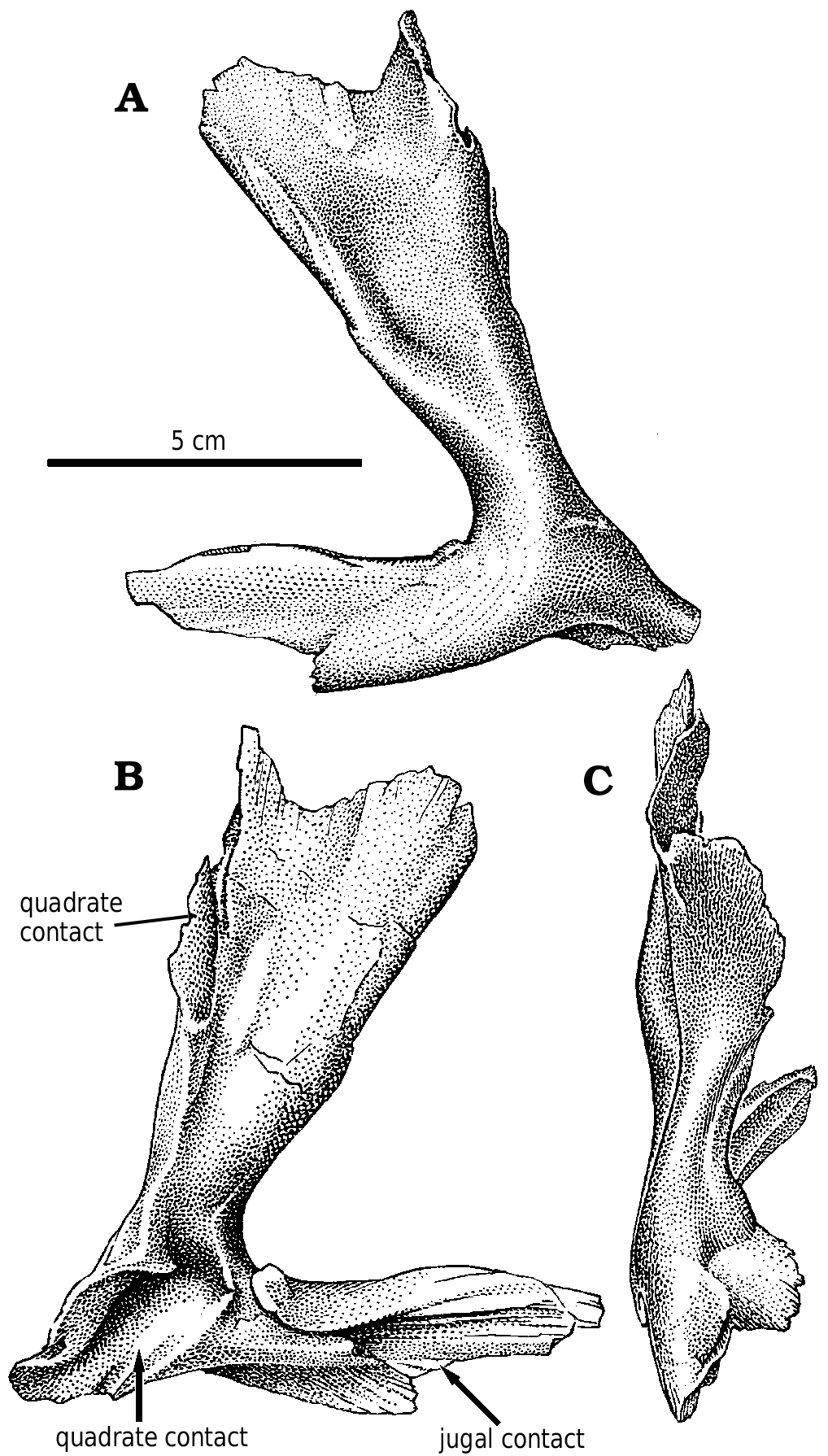
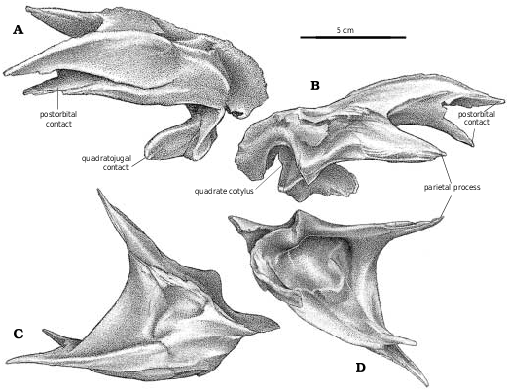
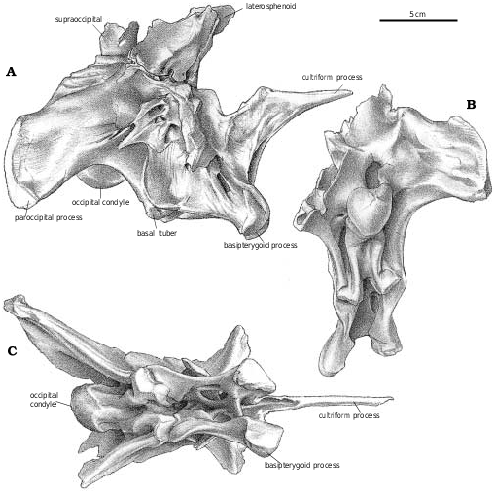
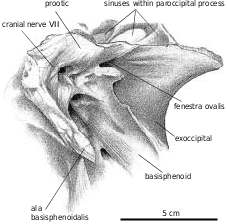
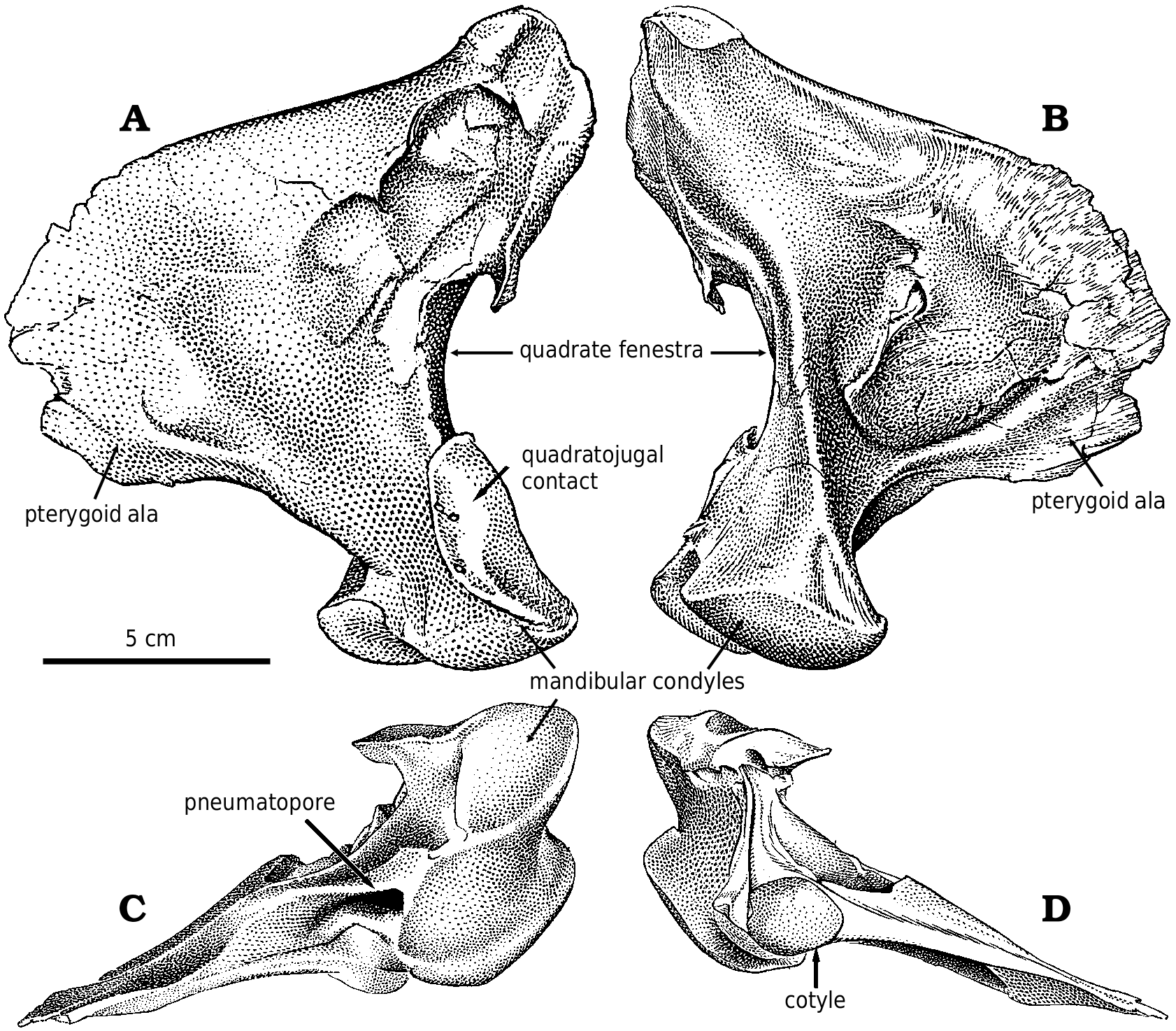

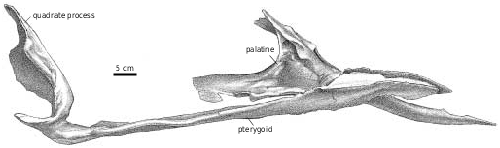
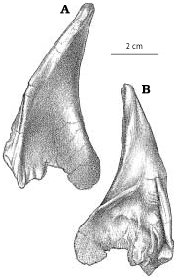
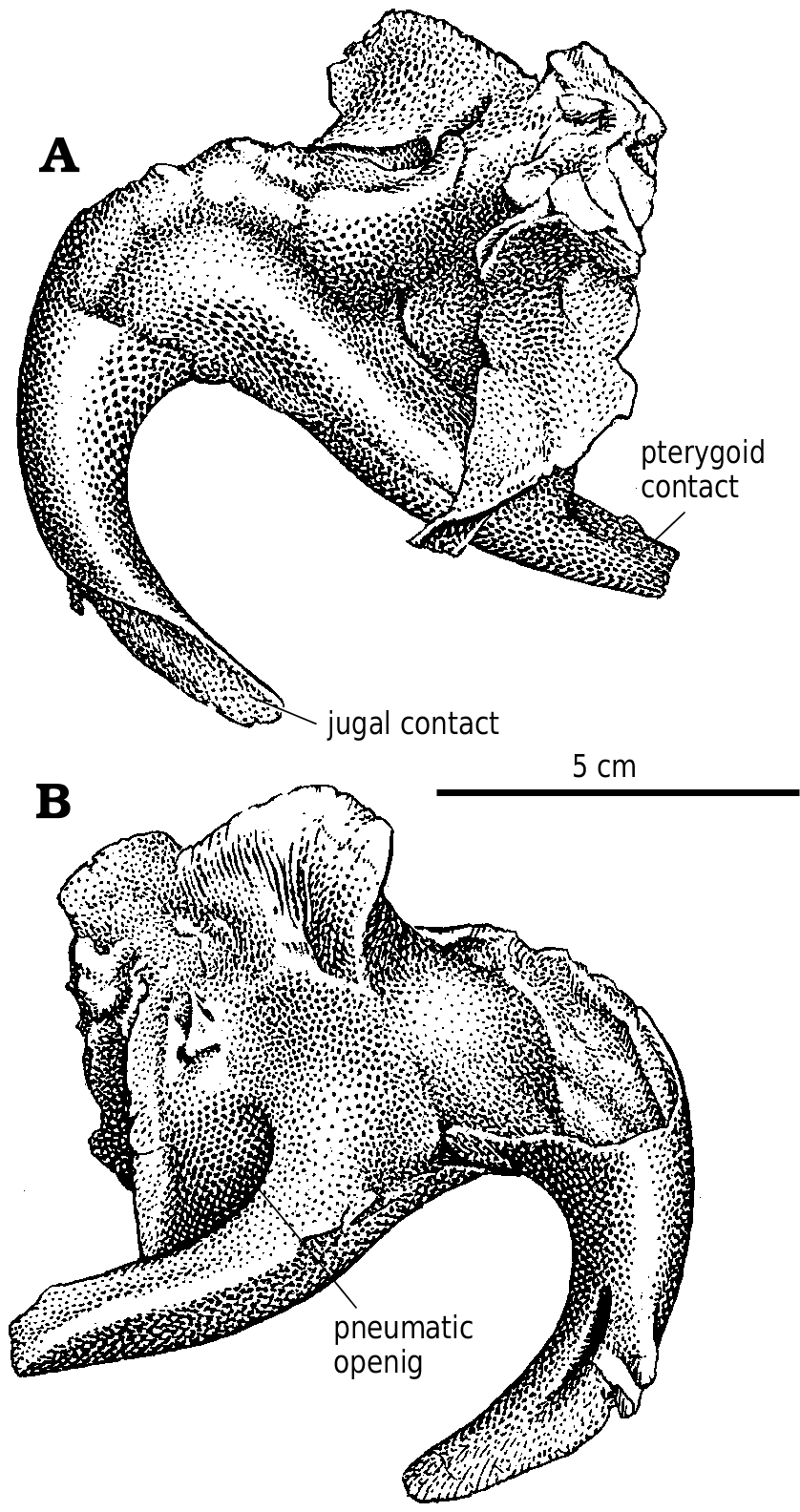
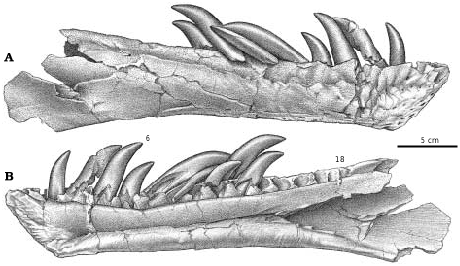
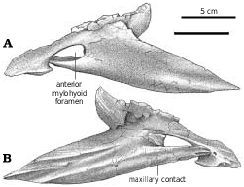

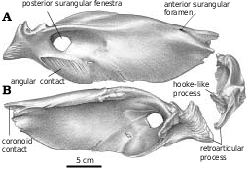
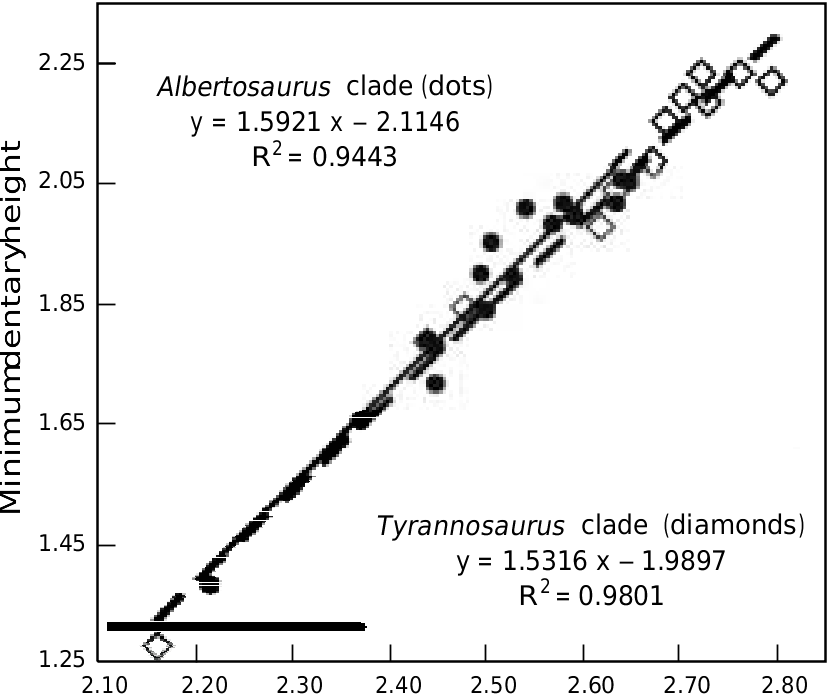
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |