
Ostarriraja parva, Marram̀ & Schult & Kriwe, 2019
|
publication ID |
https://doi.org/10.1080/14772019.2018.1486336 |
|
publication LSID |
lsid:zoobank.org:pub:8BB8F0F3-35C5-47FA-AE3C-2CBF445C4BCA |
|
DOI |
https://doi.org/10.5281/zenodo.10927191 |
|
persistent identifier |
https://treatment.plazi.org/id/B71587DF-BB7D-9D5D-2128-2D0A8E158E87 |
|
treatment provided by |
Felipe |
|
scientific name |
Ostarriraja parva |
| status |
sp. nov. |
Ostarriraja parva sp. nov.
( Figs 2 View Figure 2 , 3 View Figure 3 , 5–7 View Figure 5 View Figure 6 View Figure 7 )
1973 Dasyatis (?) sp.; Schultz in Brzobohaty & Schultz: 657, pl. 1, fig. 10.
1973 Dasyatis (?) sp.; Schultz in R ¨ogl, Schultz, & Holz: 154.
2013 Dasyatis sp. ; Schultz: 103, pl. 30, fig. 1.
Derivation of name. From the Latin word parvus meaning ‘small’, referring to the small size of the specimen; gender feminine.
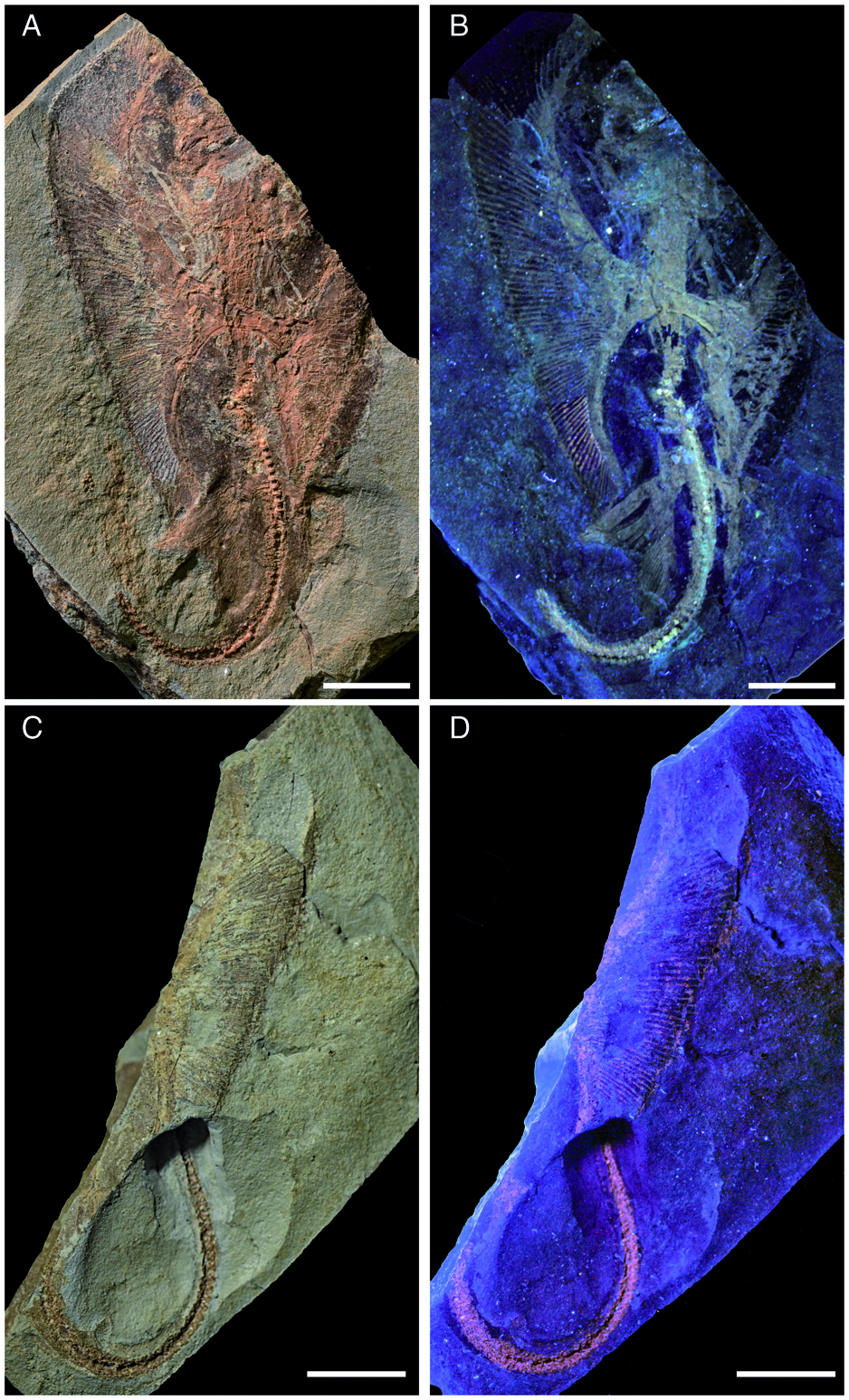
Holotype. NHMW 2005 View Materials z0283/0097, nearly complete articulated skeleton in part and counterpart preserved in a slab having length and width of 98.3 and 64.1 mm, respectively ( Fig. 2 View Figure 2 ).
Type locality and horizon. Rainbach im Innkreis, Upper Austria, Austria; Ottnangian, middle Burdigalian, early Miocene ( c. 18 Ma; see Grunert et al. 2010a).
Diagnosis. As for the genus.
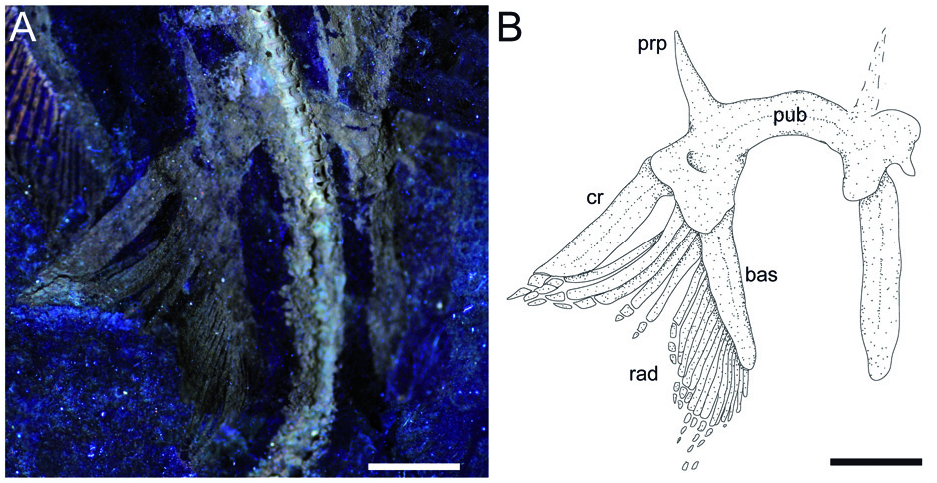
Description. Measurements and body proportions are difficult to establish since the specimen does not preserve the anterior-most portion of the body (including the rostral cartilage), the posterior-most tip of the tail, or the distal-most segments of the pectoral radials, rendering it difficult to detect the exact outline, the width of the pectoral disc and the total length of the specimen. However, the skate is quite small (possibly reaching a length of 12–14 cm in life) and is preserved as part and counterpart in a subrectangular slab having a length and width of 98.3 and 64.1 mm, respectively ( Fig. 2 View Figure 2 ). The small size of the specimen, the morphology of teeth, the poor mineralization of some skeletal structures, and the absence of claspers suggest that the specimen might represent a juvenile female skate, although the almost complete mineralization of the synarcual might indicate a late juvenile or even subadult. The specimen is displayed in ventral view, as suggested by the exposure of jaws and teeth, and the scapulocoracoid bar of the pectoral girdle lying ventrally to the vertebral axis. The body is dorso-ventrally compressed, and the disc appears largely free of dermal denticles and thorns, at least ventrally. However, multiple rows (at least three) of thorns are present along the tail, which is long and robust, but not slender and whip-like as in skates of the family Anacanthobatidae (see Hulley 1973; Last et al. 2016). Several parts of the skeleton show the typical prismatic calcification of elasmobranch fishes ( Dean & Summers 2006) and are highly visible in UV light ( Figs 2 View Figure 2 , 3 View Figure 3 , 6 View Figure 6 ). However, some of the skeletal elements, such as the distal-most segments of the pectoral radials, are not preserved, suggesting that they were still poorly or not at all mineralized. The number of dorsal fins, typically present near the extremity of the tail in skates (e.g. Last et al. 2016) is unknown in Ostarriraja , because the tip of the tail is not preserved.
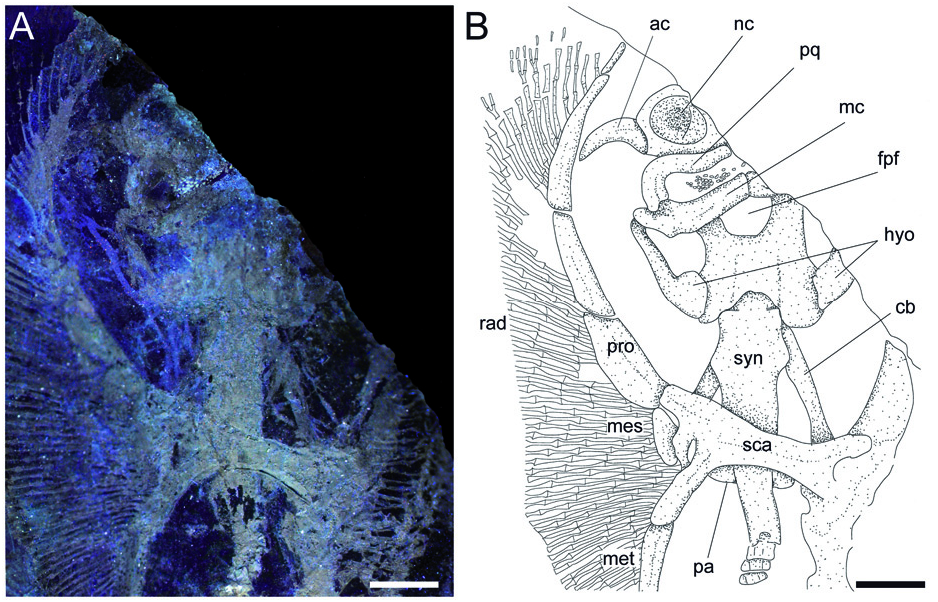
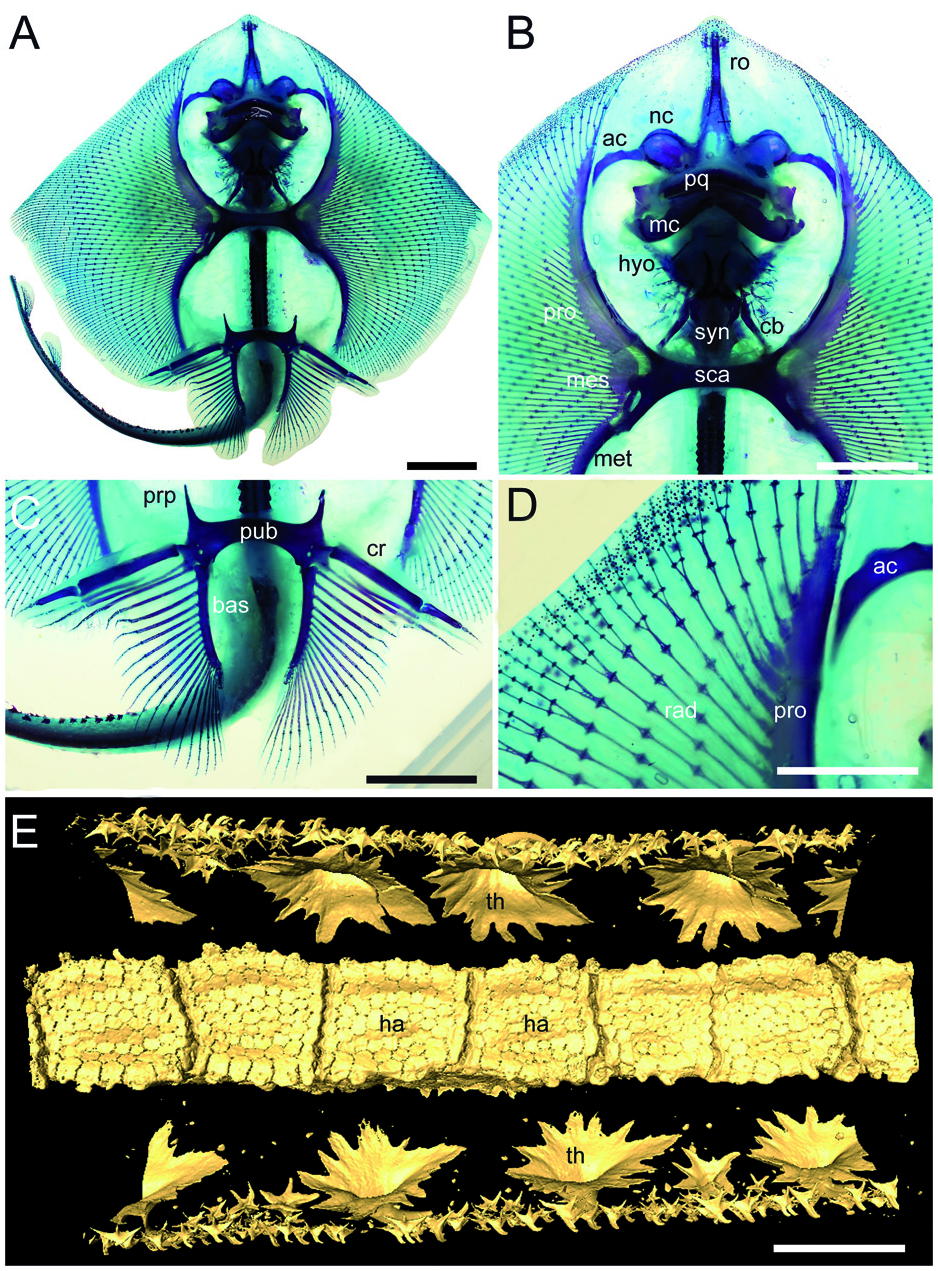
Cranium. Although the cranium is only partially preserved, it appears antero-posteriorly elongated, longer than wide, with its greatest width at the level of the nasal capsules ( Fig. 3 View Figure 3 ). The rostral cartilage is not preserved. Only one of the two nasal capsules is exposed. It appears broad and oval without the kidney-shaped basal fenestra on its antero-medial aspect typically present in Psammobatis , Irolita , Pseudoraja , Pavoraja , Notoraja , Fenestraja and Gurgesiella (see McEachran & Miyake 1990a; McEachran & Dunn 1998) but mostly resembling the condition seen in Amblyraja ( Fig. 4A, B View Figure 4 ). Since the specimen is exposed in ventral view, the presence and morphology of the preorbital processes remain ambguous (= preorbital flanges of McEachran & Miyake 1990a). The neurocranium is narrower at the level of the orbital region. The otic capsules are short and robust. Although the specimen shows the ventral side in the main slab, it is possible to recognize (possibly due to taphonomic compression) the outline of the posterior portion of the fronto-parietal fontanelle, whose posterior margin appears concave and does not display any indentation. The antorbital cartilage is massive, unbranched and arched ( Fig. 3 View Figure 3 ), and its maximum width is at the level of the articulation with the postero-lateral aspect of the nasal capsule. It tapers distally and extends laterally articulating with the third segment of the propterygium, resembling the condition of skates ( Fig. 4B View Figure 4 ).
Jaws and hyoid arch. Only the right hemi-jaws are preserved. The palatoquadrate is broadly arched, labio-lingually compressed, and slightly smaller and narrower than the Meckel’ s cartilage ( Fig. 3 View Figure 3 ). The palatoquadrate slightly tapers towards the symphysis, and possesses a strong condyle that articulates with the Meckel’ s cartilage at the mandibular articular fossa. The Meckel’ s cartilage is stouter and broader than the palatoquadrate. Its antimeres are robust, not tapering, and are separated at the symphysis. There are no labial cartilages. The hyomandibulae are narrow and elongate, enlarged and stout proximally at the articulation with the otic region of the neurocranium, and tapering distally towards their articulation with the Meckel’ s cartilage. With the exception of only the fifth pair of ceratobranchials, the ventral gill arch skeleton of Ostarriraja is not preserved. However, the fifth ceratobranchials are long and straight, and articulate with the anterior margin of scapulocoracoid.
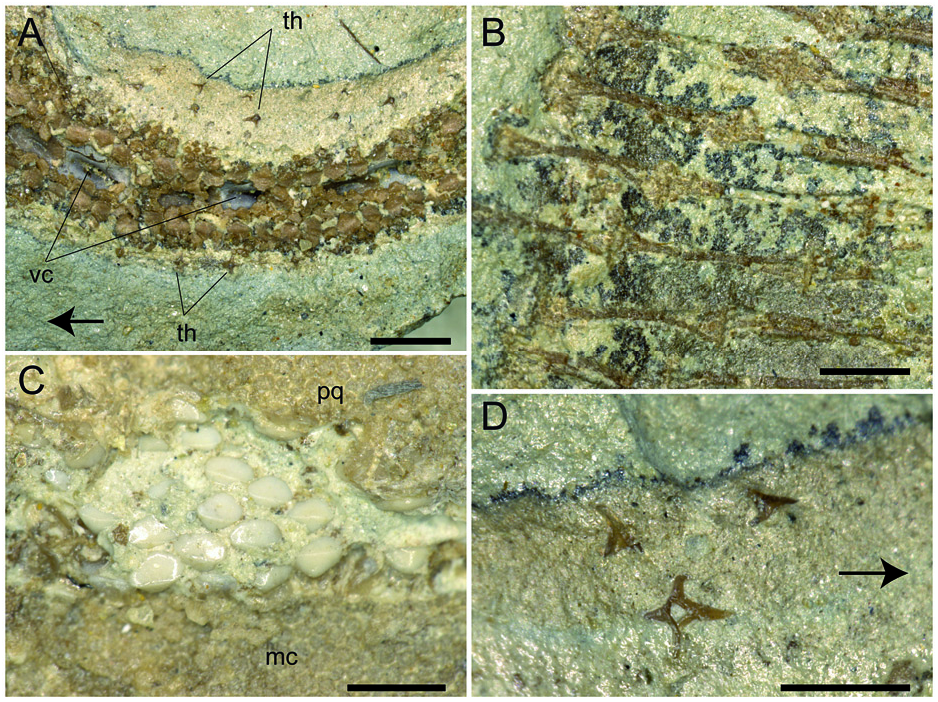
Synarcual and vertebral column. The synarcual cartilage ( Fig. 3 View Figure 3 ) is strongly calcified and tube-shaped, and its morphology is consistent with that of skates (see Claeson 2008, 2011). Anteriorly, the synarcual articulates with the occipital condyles of the chondrocranium through a subrectangular synarcual lip which rests inside the foramen magnum of the chondrocranium, a condition considered derived in skates ( Aschliman et al. 2012a). The two lateral occipital cotyles of the synarcual articulate with the occipital condyles of the chondrocranium. The synarcual extends posteriorly to the level of the shoulder girdle. The pectoral arch, formed by the fusion of suprascapulae to the dorsal median crest of the synarcual in skates ( Claeson 2008; Aschliman et al. 2012a), is partially visible posterior to the scapulocoracoid bar. However, the dorsal median crest of the synarcual and the lateral stays are not exposed in our specimen. It is not possible to detect the number of fused vertebrae that form the synarcual, or the spinal nerve foramina. Two or three unfused individual vertebral centra can be seen near the posterior margin of the synarcual. At least 13–15 trunk vertebrae (from the first distinguishable centrum to the anterior margin of the puboischiadic bar) can be recognized. About 50 vertebrae are visible from the anterior margin of the puboischiadic bar to the last portion of the tail although this number was originally higher since the distal tip of the tail is not preserved. However, it is likely that the total number of predorsal vertebrae might have been about 65–70. The vertebral centra are strongly calcified, subrectangular in shape and antero-posteriorly elongated. The vertebrae of the tail appear to be surrounded by small calcified tesserae of polygonal shape ( Fig. 5A View Figure 5 ), which probably represent the tesserae that form the prismatic calcification of the modified neural and haemal arches in modern skates ( Fig. 4E View Figure 4 ). Ribs are absent.
Pectoral fins and girdle. The scapulocoracoid is formed by a single straight and robust transverse structure, located just ventral to the synarcual arch ( Fig. 3 View Figure 3 ). The scapulocoracoid articulates anteriorly with the fifth pair of ceratobranchials. The lateral aspect of the scapulocoracoid appears to be large, and the distance between pro- and mesocondyles is less than the distance between meso- and metacondyles, contrary to some skates like Amblyraja, in which the distance between condyles is about equal ( Fig. 4B View Figure 4 ). The presence of the bridge and the postventral fenestra cannot be detected in Ostarriraja . The propterygium is long and arched, and gradually tapers distally. It is segmented and at least four segments can be recognized; the proximal one is enlarged and articulates with the procondyle on the anterior portion of the lateral margin of the scapulocoracoid. The third propterygial segment articulates mesially with the antorbital cartilage of the neurocranium. Due to the incomplete preservation of the anterior portion of the body it is not clear whether the propterygium extends anteriorly to the rostral node. The mesopterygium is small, shorter than the pro- and metapterygium. It is a single ovoid element and its external margin is more or less straight to slightly convex, apparently not fused to the radials. The metapterygium is arched and its length is less than that of the propterygium. The metapterygium gradually tapers distally, where it segments at least once. There are about 86 pectoral radials. Of these, 33 are propterygial, 10 mesopterygial and 32 metapterygial. Moreover, about 11 radials directly articulate with the scapulocoracoid between the mesopterygium and metapterygium, resembling the typical condition of skates, Pristis , panrays and guitarfishes ( Garman 1913, pls 64 and 65; Nishida 1990, fig. 32; McEachran et al. 1996, fig. 9; Aschliman et al. 2012a). Each pectoral radial is composed of at least five segments. However, since the external margin of the disc is incompletely preserved, possibly due to the incomplete mineralization of the distal-most radial cartilages in this presumably late juvenile specimen, it is possible that the number of segments was much higher. The anterior-most propterygial radials bifurcate distally at least once. The radials of Ostarriraja are calcified in chain-like patterns ( Fig. 5B View Figure 5 ), forming the so-called ‘catenated calcification’ typical of batoids with undulatory swimming mode, including skates and most of the benthic stingrays ( Schaefer & Summers 2005). Moreover, as observed in modern skates ( Fig. 4D View Figure 4 ; see also Schaefer & Summers 2005, fig. 2), the calcification is further reduced in Ostarriraja , since it consists of a single chain on the dorsal and ventral sides in all radials.
Pelvic fins and girdle. As observed in modern skates ( Fig. 4C View Figure 4 ; see also Compagno 1999; Holst & Bone 1993, fig. 1; Lucifora & Vassallo 2002, fig. 2), the pelvic fins of Ostarriraja are typically bilobed, characterized by the presence of distinct anterior and posterior lobes ( Fig. 6 View Figure 6 ). The anterior lobe is supported by a rod-like compound radial (articulated proximally to the radial condyle of pelvic girdle and distally in serial fashion with the proximal radial, which in turn is articulated to the distal radial) and three or four most anterior radials arising directly from the puboischiadic bar. The posterior lobe, conversely, is supported by the long basipterygium, articulating with the basal condyle of the pelvic girdle, which sustains 16 basipterygial radials. Pelvic girdle condyles for the compound radial and the basipterygium are therefore distinctly separated, and the pelvic fins include about 20–21 total radials each. The presence of anterior radials arising directly from the puboischiadic bar clearly distinguish the morphology of the pelvic fins of Ostarriraja from those of Cruriraja , Schroederobatis and Springeria, which are unique among skates in the absence of radials in the proximal section of the basipterygium and radials arising directly from the puboischiadic bar, therefore leaving a gap in the distribution of the pelvic-fin radials ( Bigelow & Schroeder 1948; McEachran & Miyake 1990a, fig. 12; McEachran & Dunn 1998). The puboischiadic bar is robust and wide, with a slightly convex anterior margin, although we do not exclude that taphonomy might have influenced, at least in part, the preservation of this structure, as well as the lowquality preservation of the right side of the pelvic fins. The presence of the puboischiadic foramina is difficult to detect. The prepelvic processes are moderately long, straight and pointed, extending anteriorly beyond the level of the posterior tip of metapterygia. However, the prepelvic processes are considerably shorter than one-half the width of the puboischiadic bar, distinguishing them from those of Psammobatis and Sympterygia (longer than one-half width of the puboischiadic bar) or from Schroederobatis and Springeria (long with forked tips) ( McEachran & Miyake 1990a, figs 8 and 12; McEachran & Dunn 1998). Claspers are not present, corroborating the hypothesis that NHMW 2005z0283/0097 represents a young female.
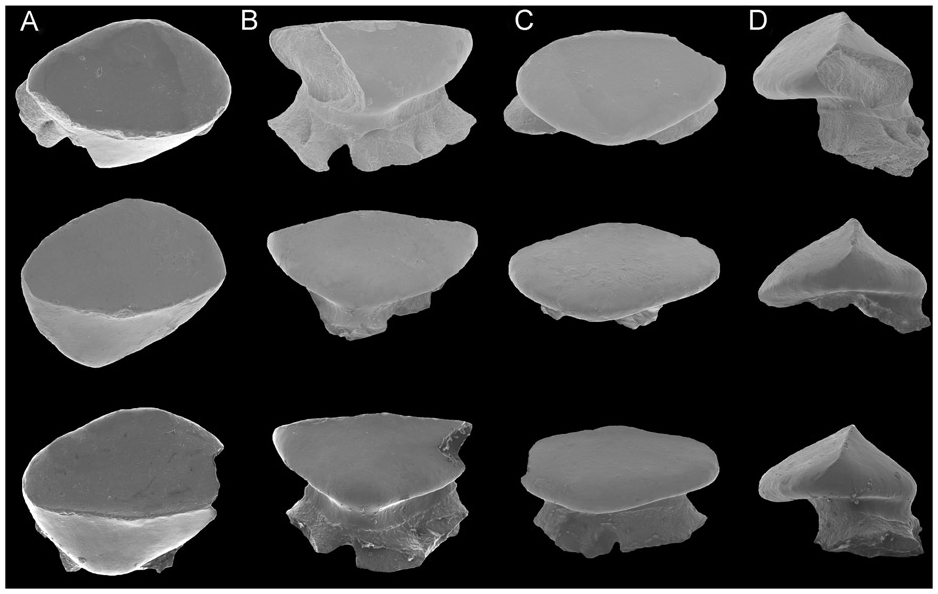
Dentition. The teeth of Ostarriraja are small ( c. 400–500 µm in crown width) and arranged in numerous rows ( Fig. 5C View Figure 5 ). The dentition is of crushing type with a gradient monognathic heterodonty. The teeth are uncusped and slightly decrease in size distally. Sexual and ontogenetic heterodonties are unknown because only a single specimen is available. The specimen shows only three or four tooth files on each jaw, resembling the condition of the early tooth development in skates (see Underwood et al. 2015).
The crown is massive, semi-oval in occlusal view, and longer mesio-distally than labio-lingually, with an arched labial edge and a lingual one with a small medial protuberance ( Fig. 7 View Figure 7 ). The teeth show a well-marked transverse cutting crest that separates the labial and lingual crown surfaces. The mesial and distal cutting edges are concavely arched and reach the margins of the crown. A labial cutting edge is absent. There is no strong cusp but the crown bears a poorly developed, semi-centrally situated and erected cone. The apron, uvula and crown ornamentation are absent. The lingual surface is weakly convex. The root is obliquely directed lingually. The root is massive, as wide as the crown but not protruding below the crown in occlusal view, relatively high and more or less oval in cross section. The root is holaulacorhize and bilobate, with unequally developed massive root lobes that widen basally to form a large base with slightly undulated margins. However, we do not exlude that the presence of unequally developed root lobes could be due to the juvenile stage of the specimen or the position of the teeth within the jaws. A pair of margino-lingual foramina is present. Their presence might indicate the presence of fusion of root lobes, although we do not exclude, again, that fusion might be related to the juvenile stage. The collar (= root coating of Herman et al. 1996) is very distinct and covers the upper part of the root stem.
Squamation. The surface of the disc of Ostarriraja appears to be largely free of dermal denticles. Thorns on the dorsal surface of the nuchal and scapular region of the disc, as well as thorns along the dorsal midline of the disc, appear absent, although this might be due to the ventral exposure of the specimen, which prevents identification of the dorsal surface of the disc. Malar and alar thorns also appear to be absent. However, it must be emphasized that malar and alar thorns are only present in mature male skates ( McEachran & Konstantinou 1996; McEachran & Dunn 1998), and their presence in Ostarriraja is very unlikely due to the sex (female) and late juvenile stage of the fossil specimen (development of denticles proceeds caudo-rostrally in skates; Miyake et al. 1999). However, multiple rows of thorns are present along the entire dorsal margin of the tail ( Fig. 5A View Figure 5 ). Thorns on the tail occur in three originally equally spaced parallel rows (one mediocaudal and two laterocaudal) formed by at least 35 similarsized and similar-shaped pungent thorns each. In dorsal view ( Fig. 5D View Figure 5 ) thorns are bilateral symmetric with four strongly developed elongated ridges forming right angles to each other, typical of the cross-type morphology of dermal denticles of some skates ( Gravendeel et al. 2002). The anterior elongate ridge is larger than the posterior one. The crown of the thorn is hook-like and obliquely implanted on the basal plate. The cusp of the crown clearly overshoots the basal plate posteriorly and the crown is almost as long as the basal plate, resembling the condition of thorns in Rajella lintea (= Dipturus linteus of Gravendeel et al. 2002).
| R |
Departamento de Geologia, Universidad de Chile |
| UV |
Departamento de Biologia de la Universidad del Valle |
| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
SuperOrder |
Batomorphii |
|
Order |
|
|
Genus |