
Coryphaenoides leptolepis Günther, 1877
|
publication ID |
https://doi.org/ 10.11646/megataxa.3.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/B711B23F-FF70-86A0-D99D-C31CFF397EA4 |
|
treatment provided by |
Plazi |
|
scientific name |
Coryphaenoides leptolepis Günther, 1877 |
| status |
|
Coryphaenoides leptolepis Günther, 1877 View in CoL
[Japanese name: Hiragashira-dara]
( Figs. 110–114 View FIGURE 110 View FIGURE 111 View FIGURE112 View FIGURE 113 View FIGURE 114 ; Table 8 View TABLE 8 ; Appendix 3-6D)
Coryphaenoides leptolepis View in CoL G̹nther, 1877:441 (original description; 4 spec., from “off the coasts of Brazil, Japan, and Mid-Pacific”, Challenger sta. 122, 237, and 246); Gilbert & Hubbs 1916:143 (listed); Nybelin 1957:268, tables V–VI (synonymy; description; NorthAtlantic; comparison with“ Co.leptolepis View in CoL ”); Iwamoto & Stein 1974:43, fig. 23 (synonymy; description); Geistdoerfer 1986:652 (North Atlantic); Geistdoerfer 1990:546 (eastern tropical Atlantic; compiled; synonymy); Iwamoto 1990:187 (in key); McAllister 1990:122 (listed; Canada); Mecklenburg et al. 2002:274 (brief description; Alaska; compiled; distribution); Moor et al. 2003:209 (New England); Mundy 2005:250 [record from Hess Seamount (not Hawaii), after Wilson & Waples (1983)].
Coryphaenoides variabilis View in CoL (not G̹nther 1878): G̹nther 1878:27 (original description; in part, 1 paralectotype, BMNH 1887.12 .7.132, Mid-Pacific, Challenger sta. 246) .
Chalinura simula Goode & Bean, 1883:199 [original description; 1 spec. from south of Nova Scotia, northwestern Atlantic, Blake sta. 308, in 1242 ftm (2271 m); 3 other specimens, juveniles, conditionally referred]; Goode & Bean 1896:412, fig. 345 (description; northwestern Atlantic); Garman 1899:395 (listed); Parr 1946:65, fig. 20 (description; illustration; western Atlantic); Nybelin 1957:264, tables V–VI, pl. III, figs. 3–4 (synonymy; description; North Atlantic; comparison with “ Ch. leptolepis View in CoL ”).
Macrurus leptolepis : G̹nther 1887:144, pl. XXXI (new combination; redescription based on lectotype, BMNH 1887.12 .7.109, from “off Pernambuco ”, Challenger sta. 122; other spec. of original type series designated as types for new species, M. liocephalus View in CoL ) .
Macrurussimulus:G̹nther 1887:145[newcombination;description after Goode & Bean (1883)]; Koefoed 1927:100 (synonymy; description; comparison with similar congeners).
Macrurus liocephalus G̹nther, 1887:145, pl. XXXVIII, fig. A. [original description; 3 spec., including BMNH 1887.12 .7.111 and BMNH 1887.12 .7.112 (also paralectotypes of Cor. leptolepis ), from “ Japan and Mid-Pacific ”, Challenger sta. 237 and 246, in 1875–2025 ftm (3429– 2750 m)] .
Macrurus armatus (not Hector 1875): G̹nther 1887:150 (description; in part, 1 paralectotype of Cor. variabilis, BMNH 1887.12.7.132, Mid-Pacific, Challenger sta. 246).
Chalinura serrula Bean, 1890:37 [original description; holotype: USNM 199612, from off Prince of Wales Island, Albatross sta. 2859, 55º20ʹN, 136º20ʹW, in 1569 ftm (2870 m); 2 paratypes collected with holotype]; Garman 1899:395 (listed).
Chalinura leptolepis: Goode & Bean 1896:414 (new combination; synonymy; description); Garman 1899:395 (listed); Marshall 1973:590, fig. 26 (lectotype designation; synonymy; description; western North Atlantic); Headrich & Merrett 1988:1332, pl. 39 (listed; North Atlantic Basin); Arruda 1997:66 (listed; Azores); Santos et al. 1997 (listed; Azores).
Chalinura liocephala : Jordan & Snyder 1901:119 (listed; Japan); Jordan & Gilbert in Jordan & Starks 1904:608 [new combination; after G̹nther (1877, 1887)]; Jordan et al. 1913:415 (listed; Japan; new Japanese name: “Hira-gashiradara”).
Coryphaenoides liocephalus: Gilbert & Hubbs 1916:143 View in CoL (new combination; listed); Okada & Matsubara 1938:449 (in key; Japan); Matsubara 1955:1309 (in key; Japan); Okamura 1970a:125 [brief description after G̹nther (1887)]; Hart 1973:225 (description; illustration; Canada); Tominaga & Uyeno 1981:489 (listed; Japan); Okamura 1984b:95, pl. 345, fig. F (compiled); Okamura 1988:95, pl. 345, fig. F (compiled); Iwamoto 1990:187 (in key); McAllister 1990:122 (listed; Canada); Nakabo 1993:362 (in key; Japan); Nakabo 2000:427 (in key; Japan); Nakabo 2002:427 (in key; Japan); Nakabo & Kai 2013:503 (in key; Japan); Motomura 2020:39 (listed; Japan).
Coryphaenoides simulus: Gilbert & Hubbs 1916:143 View in CoL (new combination; listed); Fowler 1936:457 (synonymy; description; Congo).
Coryphaenoides serrulus: Gilbert & Hubbs 1916:144 (new combination; listed).
Coryphaenoides (Coryphaenoides) liocephalus: Okamura 1970b View in CoL : table 1 (listed; Japan).
Coryphaenoides sp.: Endo et al. 2000:16, photo D (in situ observation; Nankai Trough).
Diagnosis. Pelvic-fin rays usually 9 or 10, rarely 8. Snout low, blunt, scarcely protruding beyond upper jaw. No stout tubercular scales on snout and head ridges. Uniserial row of small embedded scales along leading edges of snout, behind which a pair of naked lunate areas narrowly developed. Underside of head completely naked. Premaxillary teeth in wide tapered band, with outer series of enlarged canines and inner band of villiform teeth; mandibular teeth in uniserial row. Body scales covered with short, greatly reclined, needle-like spinules in widely spaced parallel rows; tip of last spinule in each row scarcely extending beyond posterior scale margin. Transverse scale rows below first dorsal-fin midbase 5–8. Orbit small, greatest diameter 17–21% HL. Mouth large, posterior margin of upper jaw extending to about vertical through hind rim of orbit or slightly beyond; upper-jaw length 39–46% HL; lateral corner of mouth not restricted by lip folds.Outer gill slit moderately wide, length 22–28% HL. Barbel long, slender, length 16–25% HL. Interdorsal space greater than first dorsal-fin base length. Height of first dorsal fin less than HL (64–75% HL); second spinous ray serrated along its leading edge; first dorsal-fin rays II,8–10. Origin of second dorsal fin posterior to that of anal fin. Outer pelvic-fin ray greatly prolonged, its tip extending well beyond anal-fin origin. Body generally pale, but posterior portion of tail much darker.
Material examined. 45 specimens. Lectotype of Coryphaenoides leptolepis: BMNH 1887.12.7.109 (91.6 mm HL, 440+ mm TL), off Pernambuco, Brazil, southwestern Pacific, 9.0833ºS, 34.8333ºW, 350 ftm (641 m), Challenger sta. 122, trawl, 10 Sept. 1873. Paralectotypes of Co. leptolepis: BMNH 1887.12.7.111 (1, 84.5 mm HL, 406+ mm TL; also lectotype of Macrurus liocephalus , designated herein), off Boso Peninsula, Chiba Pref., Japan, 34.6167ºN, 140.5333ºE, 1875 ftm (3429 m), Challenger sta. 237, trawl, 17 Jun. 1875. BMNH 1887.12.7.112 (1, 36.0 mm HL, 198 mm TL; also paralectotype of M. liocephalus , designated herein), east of Emperor Seamounts, Mid-Pacific, 36.1667ºN, 178.0000ºE, 2050 ftm (3750 m), Challenger sta. 246, trawl, 2 Jul. 1875. Holotype of Chalinura serrula: USNM 45358 (57.5 mm HL, 283+ mm TL), off Prince of Wales Island, Canada, Gulf of Alaska, northeastern Pacific, 55.3667ºN, 136.3333ºW, 1569 ftm (2870 m), Albatross sta. 2859, large beam trawl, 28 Aug. 1888. Paratypes of Ch. serrula: USNM 199612 (2, 34.2–38.3 mm HL, 146+–198+ mm TL), collected with holotype. Holotype of Chalinura simula : * MCZ 25824 (458 mm TL, fide Goode & Bean 1883:199; examination based on only photographs; Fig. 111F View FIGURE 111 ), south of Nova Scotia, off northeastern extremity of George’s Bank, northwestern Atlantic, 41.4117ºN, 65.5917ºW, 1242 ftm (2272 m), R/V Blake, sta. 308, 29 Jun. 1880 (examination based on only photographs). Paralectotype of Coryphaenoides variabilis: BMNH 1887.12.7.132 (1, 36.1 mm HL, 179+ mm TL), east of Emperor Seamounts, Mid-Pacific, 36.1667ºN, 178.0000ºE, 2050 ftm (3750 m), Challenger sta. 246, trawl, 2 Jul. 1875. Non-types: Japan: BSKU 96940 (1, 63.2 mm HL, 330+ mm TL), BSKU 96941 (1, 71.2 mm HL, 292+ mm TL), BSKU 96942 (1, 66.0 mm HL, 355+ mm TL), BSKU 96943 (1, 71.2 mm HL, 377 mm TL), BSKU 96944 (1, 60.3 mm HL, 313 mm TL), BSKU 96945 (1, 56.9 mm HL, 292 mm TL), BSKU 96946 (1, 56.8 mm HL, 297+ mm TL), southeast of Cape Erimomisaki, Japan Trench, 41.2353ºN, 144.0357ºE, 2977–3014 m, R/V Tansei-maru, cr. KT-93-15, sta. M7, 3-m beam trawl, coll. Y. Machida, 17 Nov. 1993; BSKU 96473 (1, 95.1 mm HL, 509 mm TL), off Kinkazan, 38.4748ºN, 143.3470ºE, 2698–2814 m, R/V Tansei-maru, cr. KT-08-27, sta. K-3, 3-m ORE beam trawl, coll. H. Endo and N. Nakayama, 23 Oct. 2008; NSMT-P 97650 (3, 60.8–84.9 mm HL, 306+– 444 mm TL), off Kesennuma, Japan Trench, 38.7983ºN, 144.1345ºE, 7340–7433 m, R/ V -Hakuho-maru, cr. KH-01-02, sta. TD-7, 30 Sept. 2001. Australia: FMNH 5777 (1, 39.3 mm HL, 137+ mm TL), south of Western Australia, 50.0167ºS, 123.0667ºE, 1800 ftm (3292 m), Challenger sta. 158, trawl, 7 Mar. 1874. Northeastern Pacific: FMNH 117120 (2, 28.9–47.7 mm HL, 141+–221+ mm TL), off Newport, 44.7658ºN, 125.5294ºW, 1750 m, R/V Wecoma, cr.W9704A, sta. 5, 40- ft otter trawl, coll. J. Voightt, 17 Apr. 1997; FMNH 117117 (1, 49.5 mm HL, 248+ mm TL), off Newport, 44.7581ºN, 125.5361ºW, 2850 m, R/V Wecoma, cr. W9704A, sta. 1, 40-ft otter trawl, coll. J. Voightt, 16 Apr. 1997; * CMNFI 1965-0321.3 (1, 84 mm HL, 342+ mm TL; examination based on only photographs provided by S. Tessier), south of Moresby Island, 51.0500ºN, 132.2200ºW, 2762 m, 8 Aug. 1965. North Atlantic: ZMUB 3384 (1 of 2, 85.8 mm HL, 315+ mm TL), 42.4333ºN, 25.7500ºW, 3120 m, R/V Michael Sars, sta. 88, 18 Jul. 1910; ZMUB 16803 (1, 62.4 mm HL, 326 mm TL), Mid-Atlantic Ridge, 42.9167ºN, 30.3333ºW, 2660–2670 m, R/V G.O. Sars, cr. MAR-ECO, sta. 1149, 7 Jul. 2004; ZMUB 16805 (1, 64.0 mm HL, 325+ mm TL), Mid-Atlantic Ridge, 53.0333ºN, 33.6000ºW, 2955–3071 m, R/V G.O. Sars, cr. MAR-ECO, sta. 1165, 24 Jul. 2004; * ZMUB 17836 (1, 71.0 mm HL, 352+ mm TL), Mid-Atlantic Ridge, 51.3297ºN, 28.8690ºW, 3505–3527 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 54, local sta. 377, 16 Jul. 2004; ZMUB 17838 (1, 71.4 mm HL, 344+ mm TL), * ZMUB 17839 (1, 124+ mm TL, poor condition), * ZMUB 19319 (2, 74.7–75.3 mm HL, 378+– 456 mm TL), Mid-Atlantic Ridge, 51.5493ºN, 30.9683ºW, 3452–3465 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 64, local sta. 381, 21 Jul. 2004; ZMUB 17840 (1, 77.1 mm HL), ZMUB 19517 (5, 83.3–105 mm HL, 390+– 578 mm TL), Mid-Atlantic Ridge, 51.0343ºN, 33.6117ºW, 2995–3071 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 66, local sta. 383, 24 Jul. 2004; * ZMUB 19287 (1, 89.5 mm HL, 467+ mm TL), ZMUB 19288 (1, 107 mm HL, 567 mm TL), Mid-Atlantic Ridge, 42.9220ºN, 28.1392ºW, 2973–2979 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 52, local sta. 374, 13 Jul. 2004; * ZMUB 19338 (1, 88.6 mm HL, 452+ mm TL), Mid-Atlantic Ridge, 53.1387ºN, 34.7660ºW, 2306–2374 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 68, local sta. 384, 25 Jul. 2004; * ZMUB 17837 (1, 100+ mm TL, poor condition), Mid-Atlantic Ridge, 51.3297ºN, 28.8690ºW, 3505–3527 m, R/V G.O. Sars, cr. MAR-ECO, supersta. 54, local sta. 377, 16 Jul. 2004; ZMUC P372974 (1, 67.0 mm HL, 330+ mm TL), ZMUC P372975 (1, 91.3 mm HL, 416+ mm TL), northeastern Atlantic, 49.9167ºN, 13.4333ºW, Challenger sta. 51804, 26 Sept. 1983. Uncertain variant ( Japan): BSKU 96947 (1, 80.9 mm HL, 384+ mm TL, uncertain variant), southeast of Cape Erimomisaki, Japan Trench, 40.7270ºN, 143.7097ºE, 2607–2609 m, R/V Tansei-maru, cr. KT-93- 15, sta. M6, 3-m beam trawl, coll. Y. Machida, 18 Nov. 1993.
Redescription. General features are shown in Figs. 110–111 View FIGURE 110 View FIGURE 111 . Counts and measurements are given in Table 8 View TABLE 8 . Body deepest at first dorsal-fin origin, gradually tapering to long tail. Trunk short, moderately compressed, width over pectoral-fin bases 1.4–1.7 in depth at first dorsal-fin origin. Head large, HL about 4.8–5.2 in TL; head bones moderately stout; flesh firm. In the largest specimen (BSKU 96473, 80.9 mm HL), dorsal contour of nape moderately humped, forming distinct depression above orbit. Snout short, low, barely protruding beyond upper jaw, length 1.2–1.4 times as long as orbit diameter; ventral contour almost vertical in lateral view.Orbit small, greatest diameter 2.9–3.5 in postorbital length. Interorbital space slightly concave in preserved specimens, width 0.7–0.8 in orbit diameter.Mouth large, subterminal, upper-jaw length 0.4–0.5 in orbit diameter; posterior margin of maxilla reaching vertical through hind rim of orbit or slightly beyond; lateral corner of mouth not restricted by lip folds; lips thin, not papillose. Suborbital region almost flat, lacking stout bony ridge. Preopercle large, hind margin almost vertical, forming broad lobe posteroventrally; preopercular ridge with prominent backward extension at angle. Interopercle mostly covered by preopercle, but posterior portion exposed as narrow flap. Hind margin of subopercle concave, smoothly joined with opercle. Gill membranes narrowly joined across isthmus, with broad posterior free fold; gill opening wide, extending forward to below posterior margin of lower jaw. Outermost gill slit restricted by skin folds but moderately wide, length 0.7– 0.9 in orbit diameter. Outer gill rakers on first arch small, flat, lappet-like; other rakers moderately high, ridgelike; all rakers armed with short, slender spines distally; no rakers on inner side of fourth arch. Gill filaments moderately developed. Barbel long, slender throughout its length, 0.9–1.1 in orbit diameter.
Anus immediately anterior to anal-fin origin; periproct very narrow. Ventral light organ absent. Pyloric caeca 10 (based on lectotype of C. liocephalus ).
Premaxillary teeth in tapered band, with wide villiform band of small conical teeth (about 8 rows near symphysis) and outer row of widely spaced, enlarged canines; posterior margin of tooth band falling short of lateral corner of mouth. Mandibular teeth slightly smaller than outer teeth of premaxillary, arranged in uniserial row, but rarely clustered near symphysis.
Body scales moderately small, highly deciduous, completely missing in most of the specimens examined. In 71.2 mm HL specimen (BSKU 96941), those on dorsum below interdorsal space covered with short, greatly reclined, needle-like spinules in 7 parallel rows ( Fig. 112A View FIGURE112 ); tip of last spinule in each row scarcely extending beyond posterior scale margin; no reticulate structures on unexposed potions. Body entirely and uniformly scaled except for fins.
TABLE 8. (Continued)
TABLE 8. (Continued)
No prominent enlarged scutes on tip and lateral angles of snout. Head ridges lacking stout modified scales. Single row of small embedded scales along leading edge of snout, behind which a pair of naked lunate areas narrowly developed. Ventral surfaces of head including snout, lower half of suborbital area, mandibular rami, lower preopercle, and exposed portion of interopercle completely naked. Nasal fossa also lacking scales.
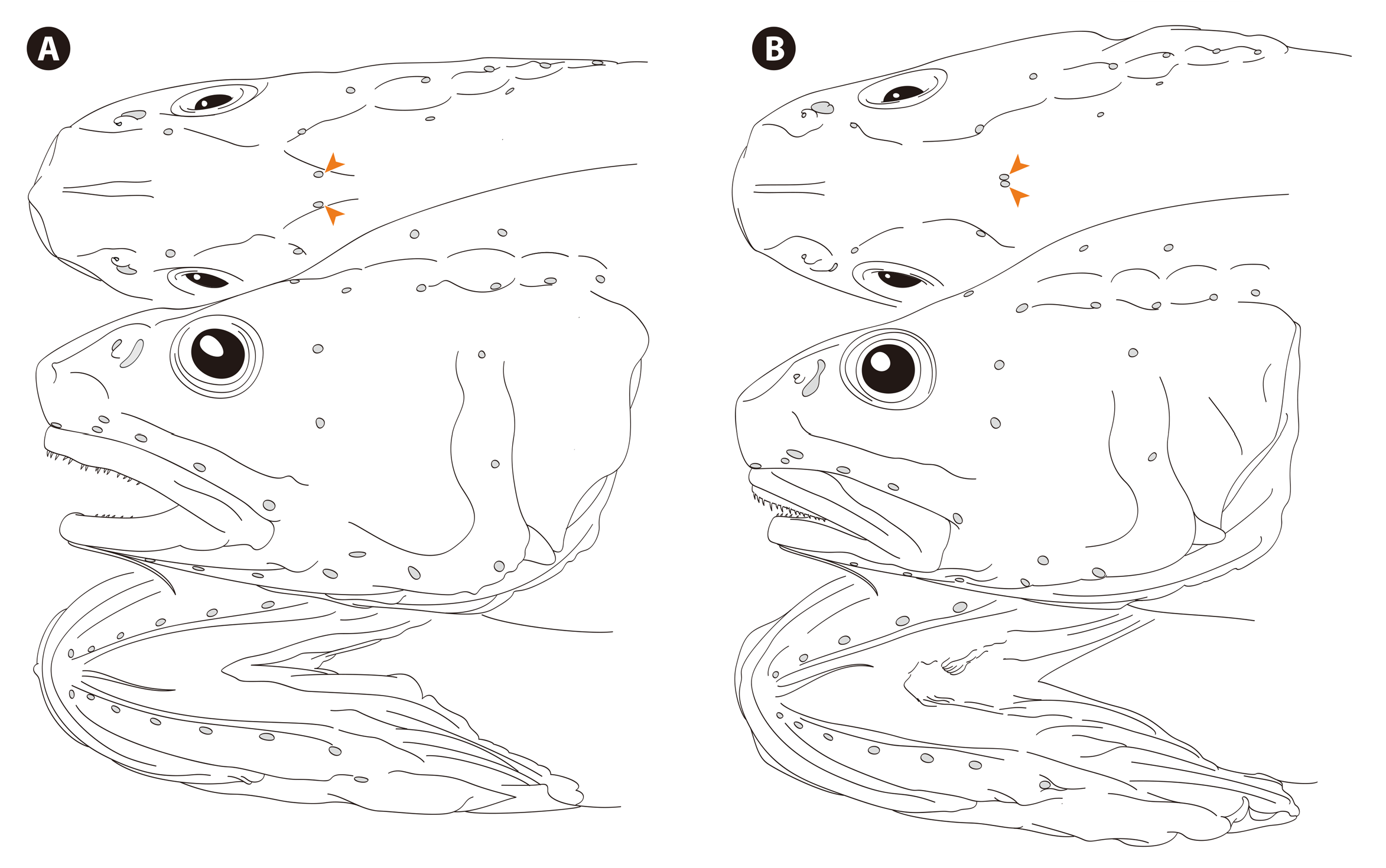
Cephalic sensory canals well developed, broad, slightly inflated when fresh ( Fig. 110 View FIGURE 110 ). Open pores present and serially arranged along all canals ( Fig. 113 View FIGURE 113 ); those on infraorbital, preopercular, and mandibular canals enlarged, easily discernible even in preserved specimens.
Origin of first dorsal fin slightly posterior to vertical through pectoral-fin base; first dorsal fin well developed, height 2.6–2.8 times as long as its base length; second spinous ray not especially elongate, armed along its leading edge with short, greatly reclined, needle-like denticles. Second dorsal fin well separated from first dorsal, interdorsal space 1.3–1.9 times as long as first dorsal-fin base length; the fin originating above base of 8th–10th anal-fin rays. Pelvic-fin base slightly anterior to vertical through pectoral-fin base; outermost ray greatly prolonged, its tip reaching bases of 5th–10th anal-fin rays, when laid back.
Color when fresh ( Fig. 110 View FIGURE 110 ). Ground color of head and body pale pinkish gray, gradually becoming darker posteriorly; abdomen bluish, especially when young; upper jaw and gill cover prominently darker; gular and branchiostegal membranes deep blue, but posterior margins of branchiostegal membranes narrowly edged with black; first dorsal, pectoral, and pelvic fins pale, but free portion of outer pelvic-fin ray blackish; anal fin uniformly dark dusky.
Color in alcohol ( Fig. 111 View FIGURE 111 ). Dorsum trunk light beige, abdomen dark blue, tail gradually becoming darker posteriorly; head generally pallid, but opercle distinctly dark; orbit narrowly edged in black; lips blackish; septum between nostrils dark; oral and gill cavities dull black; gill rakers and arches dusky, filaments yellowish brown; chin dark, but gular membrane paler; branchiostegal membranes mostly pale, but abruptly blackish along their margins; chin barbel darker basally, paler distally; first dorsal fin pale to dusky overall, but tip of second spinous ray slightly blackish; second dorsal and anal fins mostly blackish, paler anteriorly; pectoral and pelvic fins generally pale, but uppermost pectoral-fin ray and outermost pelvic-fin ray prominently dark.
Size. The maximum size is about 58 cm TL (ZMUB 19517, 578 mm TL, Mid-Atlantic Ridge).
Variation. In juveniles, small but prominent black melanophores sparsely scattered over the body surface; this characteristic pigmentation disappears in large adults.
Development. Larval morphology was described by Stein (1980) and Merrett (1986).
Distribution. Widespread from the continental slope to hadal depths of the world’s oceans in 640–7433 m (Marshall 1973; Iwamoto & Stein 1974; Mecklenburg et al. 2002; this study). In Japan, known only from the Pacific off Honshu and Hokkaido at depths of 2698‾ 7433 m (Appendix 3-6D). Rare in its depth range.
Remarks. Coryphaenoides leptolepis was originally described by G̹nther (1877) based on specimens collected from Brazil, Japan, and the Mid-Pacific. A decade later, G̹nther (1887) provided a further description of this species (as Macrurus leptolepis ) based on only the Brazilian syntype (from Challenger sta. 122; Fig. 111A View FIGURE 111 ), allocating the remaining type specimens from Japan (1 spec. from sta. 237; Fig. 111B View FIGURE 111 ) and the Mid-Pacific (2 spec. from sta. 246; Fig. 111C View FIGURE 111 ) to his new species C. liocephalus (described as M. liocephalus based on these three specimens).According to G̹nther (1887: annotation on p. 144), C. liocephalus differs from C. leptolepis by having a “shorter snout”, while he did not provide any other characters to separate the two species.
The synonymy between C. leptolepis and C. liocephalus has been uncertain over the past several decades ( Iwamoto & Stein 1974; Iwamoto 1990; Mecklenburg et al. 2002). Although C. leptolepis was widely recorded from the Northern Hemisphere of the Atlantic and eastern Pacific, no additional specimens were collected from the type locality (off Brazil). By contrast, C. liocephalus was represented by only its type specimens, except for a record from off British Columbia by Hart (1973). Due to a lack of additional materials of C. liocephalus , previous authors did not attempt to confirm the validity of this species, and they tentatively considered C. liocephalus valid (see the Nomenclatural discussion). A close similarity between the two nominal species was first suggested by Iwamoto & Stein (1974), who wrote that “ C. liocephalus appears extremely close to, and may be conspecific with, C. leptolepis ”. The difficulty of separating these species was also indicated by Iwamoto (1990); in his key to species of the genus ( Iwamoto 1990:187–188), C. leptolepis keys out together with C. liocephalus at 18a and 21b, but no further couplets are provided to distinguish the two species. Citing a personal communication from Iwamoto (31 Aug. 1999), Mecklenburg et al. (2002:270) indicated that the tissue samples from their type localities could help determine their taxonomic status, and if significant differences exist, specimens called C. leptolepis from the North Pacific correspond with C. liocephalus , and C. leptolepis would be restricted to the Atlantic.
Furthermore, C. leptolepis has two other junior synonyms, viz., C. simulus ( Goode & Bean, 1883) and C. serrula (Bean, 1980) . Six years after G̹nther (1877), Goode & Bean (1883) described C. simulus (as Chalinura simula ) based on a single specimen collected from the south of Nova Scotia, northwestern Atlantic (Blake sta. 308) ( Fig. 111F View FIGURE 111 ). Although this species was initially considered to be most similar to C. affinis G̹nther, 1878, Koefoed (1927:102) subsequently recognized its close similarity with C. leptolepis , suggesting their differences to be “extremely small and uncertain”. Nybelin (1957:268), who re-examined the type of C. leptolepis , distinguished these two species by several morphometric characters, including the isthmus–anus distance, preanal length, predorsal length, and barbel length. However, the suggested differences were quite subtle and based on a limited number of specimens, viz., only two specimens for C. simulus combined with data given by Koefoed (1927; 7 spec.) and Parr (1946; 1 spec.). Marshall (1970:591) indicated that the differences in proportions are likely due to dehydration of the type of C. leptolepis , or simply attributable to intraspecific variation, and C. simulus was therefore regarded as a junior synonym of C. leptolepis . Iwamoto & Stein (1974) further confirmed the two nominal species to represent a single species, showing greater variation for the morphometric characters based on additional specimens from the northeastern Pacific (16 spec.) and northwestern Atlantic (5 spec.).
The second synonymized species, C. serrula , was originally described by Bean (1890; as Chalinura serrula ) based on three specimens collected in the Gulf of Alaska (Albatross sta. 2859) ( Fig. 111E View FIGURE 111 ). Koefoed (1927:103) distinguished this species from C. simulus (= C. leptolepis ) by the combination of the orbit diameter and the number of first dorsal-fin rays. However, Iwamoto & Stein (1974) showed complete overlap in these characters based on specimens from the northeastern Pacific and northwestern Atlantic, and thus C. simulus was regarded as a junior synonym of C. leptolepis .
During the KT-08-27 cruise of the R/V Tansei-maru, the author collected a single specimen of Coryphaenoides from off Kinkazan, Miyagi Pref., Japan, at a depth of 2698–2814 m ( Fig. 110 View FIGURE 110 B–D). This specimen was then identified as C. liocephalus , but a further examination revealed that it can also be referable to C. leptolepis . In the course of this study, 11 additional Japanese specimens of the same species were found among the collections of BSKU and NSMT. The type series of C. leptolepis and C. liocephalus were examined to elucidate their taxonomic relationship, along with the new materials from Japan and non-type specimens of C. leptolepis from both the Pacific and Atlantic. The type series of C. simulus and C. serrula were also examined to reconfirm their conspecificity with C. leptolepis .
Comments on type specimens. As noted in the last section, C. leptolepis was originally described from four specimens collected from Brazil (1 spec., BMNH 1887.12.7.109, 91.6 mm HL, 440+ mm TL, Fig. 111A View FIGURE 111 ), Japan (1 spec., BMNH 1887.12.7.111, 84.5 mm HL, 406+ mm TL, Fig. 111B View FIGURE 111 ), and the Mid-Pacific (2 spec., including BMNH 1887.12.7.112, 36.0 mm HL, 198 mm TL, Fig. 111C View FIGURE 111 ). However, G̹nther (1887:144) subsequently restricted this species to Brazil, allocating the other three specimens from Japan and the Mid-Pacific to the type series of C. liocephalus . G̹nther’s (1887) act is not considered to be a lectotype designation for C. leptolepis ; the lectotype was designated by Marshall (1973:590), who recognized the Brazilian specimen as “the type” of this species [see also Eschmeyer (1998), Art. 74.5 of the ICZN (1999), and Fricke et al. (2020)]. In accordance with Marshall’s statement, the type specimens of C. liocephalus are also regarded as the paralectotypes of C. leptolepis . To fix the scientific name of C. liocephalus, BMNH 1887.12.7.111 from Japan is herein designated as the lectotype of this species, and BMNH 1887.12.7.112 from the Mid-Pacific automatically becomes a paralectotype. Unfortunately, one of the paralectotypes of C. liocephalus (and also C. leptolepis ) from the Mid-Pacific was not found in the course of this study.
According to Eschmeyer (1998) and Fricke et al. (2020), BMNH 1887.12.7.113 (not discovered during this study) is not only a paralectotype of C. leptolepis , but also a syntype of C. murrayi G̹nther, 1878. This specimen was collected from New Zealand (Challenger sta. 168), and is obviously not a type specimen of C. leptolepis . In the original description of C. murrayi , G̹nther (1878) did not indicate how many specimens were involved in his description, nor designate a holotype of this species. According to G̹nther (1878:146), five specimens were collected from the type locality, and these specimens constitute the type series of C. murrayi (see Eschmeyer 1998; Fricke et al. 2020). Iwamoto & Shcherbachev (1991) considered BMNH 1887.12.7.113 as the “putative holotype ”, although they recognized that the type series includes multiple specimens. Despite that the use of the term “ holotype ” was wrong, Iwamoto & Shcherbachev’s statement would be regarded as a lectotype designation for C. murrayi , based on Art. 74.5 of the ICZN (1999).
According to Eschmeyer (1998) and Fricke et al. (2020), BMNH 1887.12.7.132 (36.1 mm HL, 179+ mm TL, Fig. 111D View FIGURE 111 ) has the following type status: paralectotype of C. leptolepis ; syntype of C. liocephalus ; and paralectotype of C. variabilis . This specimen is only a paralectotype of C. variabilis , and is not included in the type series of either C. leptolepis or C. liocephalus . This specimen was re-identified as C. leptolepis by Wilson & Waples (1983:1136), and their identification was reconfirmed in this study.
Nomenclatural discussion. The present examination of the type series of C. leptolepis and C. liocephalus , along with additional specimens from the Pacific and Atlantic, revealed that the two species are not separable morphologically. In the “ Challenger ” Report, G̹nther (1887: annotation on p. 144) distinguished the two species only by the snout length, with C. liocephalus having a “shorter snout”. However, it was revealed here that their type specimens cannot be distinguished by the snout length (26% HL in C. leptolepis and 24–26% in C. liocephalus ; Table 8 View TABLE 8 ). According to Iwamoto & Stein (1974:45, table 1), modal counts of first dorsal- and pectoral-fin rays are slightly different between the Pacific and Atlantic populations of C. leptolepis (modally 19 vs. 21). However, the differences are negligible when the additional specimens examined are included in the analysis.Based on G̹nther’s(1887)descriptions, Iwamoto & Stein (1974:45) implied that C. leptolepis might be distinguished from C. liocephalus by its paler color (“dirty whitish” vs. “blackish”). However, this difference appears to be attributable to intraspecific geographical variation. Other characters, including dentition, squamation, and fin position, are almost identical between the Atlantic and Pacific populations, and C. liocephalus is herein synonymized with C. leptolepis .
Similarly, C. serrula was reconfirmed as a junior synonym of C. leptolepis in this study. The holotype of C. serrula ( Fig. 111E View FIGURE 111 ) agrees well with the lectotype of C. leptolepis ( Fig. 111A View FIGURE 111 ), with no substantial differences in morphological characters. Counts and measurements of the type series of C. serrula also lie within the range of variation for C. leptolepis ( Table 8 View TABLE 8 ). These two species cannot be distinguished morphologically, and therefore represent a single species.
Regarding C. simulus , this study follows Marshall (1973) and Iwamoto & Stein (1974) in regarding the species as a junior synonym of C. leptolepis . Although the holotype of C. simulus was not directly examined in this study, its photographs ( Fig. 111F View FIGURE 111 ) were kindly provided by Karsten E. Hartel, the former collection manager of the Museum of Comparative Zoology of Harvard University (MCZ). This specimen is now in poor condition, with the head skin badly deteriorated, all fins heavily damaged, and the posterior half of the tail separated from the rest of the body. Unfortunately, the original description given by Goode & Bean (1883) is too brief to determine the taxonomic status of this species, without any illustrations of its morphology. However, the general appearance of the holotype of C. simulus agrees well with what we regard as C. leptolepis , and there seems little doubt of their conspecificity.
Relationships. Coryphaenoides leptolepis was previously considered to belong to the subgenus Chalinura Goode & Bean, 1883 , whose type species is C. simulus (junior synonym of C. leptolepis ; see the Nomenclatural discussion). Although originally established as a distinct genus, most recent authors relegated Chalinura to a subgenus of Coryphaenoides (e.g., Iwamoto & Stein 1974; Iwamoto & Sazonov 1988; Iwamoto 1990; Iwamoto & Shcherbachev 1991). This group has been widely recognized as a valid taxon of grenadiers, but the name is preoccupied by Chalinura Dalman, 1826 of the spider family Hersillidae (Arachnida) (see also Dunlop & Jekel 2008; Fricke et al. 2020). A replacement name, Fuyangia , has been provided by Whitley (1931) for the Pisces Chalinura .
According to Iwamoto (1990) and Iwamoto & Shcherbachev (1991), this group comprises the following 12 species: C. brevibarbis ( Goode & Bean, 1896) restricted in the North Atlantic; C. fernandezianus (G̹nther, 1887) known only from off Juan Fernandez Island, southeastern Pacific; C. grahami Iwamoto & Shcherbachev, 1991 distributed in the Indian Ocean, the southeastern Atlantic off South Africa, and the southwestern Pacific off Australia; C. leptolepis ; C. liocephalus ; C. mcmillani Iwamoto & Shcherbachev, 1991 widespread in the Southern Hemisphere, including the southeastern Atlantic, western and central Indian Ocean, southern Australia, and New Zealand; C. mediterraneus ( Giglioli, 1893) sporadically recorded from the North Atlantic; C. murrayi G̹nther, 1878 known from the western Indian Ocean, southern Australia, New Zealand, and Fiji; C. profundicola ( Nybelin, 1957) known only from the northeastern Atlantic; C. serrulatus G̹nther, 1878 widely distributed in the Indian Ocean and southwestern Pacific; and C. striatura Barnard, 1925 and C. subserrulatus Makushok, 1976 widespread in the Southern Hemisphere. Coryphaenoides liocephalus is synonymized with C. leptolepis in this study, which reduces the total number of species to 11. On the other hand, Iwamoto & Shcherbachev (1991) recognized two subspecies of C. serrulatus : C. s. oceanus Iwamoto & Shcherbachev, 1991 from the central and western Indian Ocean; and C. s. subserrulatus from New Zealand and Australia. Iwamoto & Graham (2001) also reported two juveniles collected from off the east coast of Australia under the name Coryphaenoides sp. cf. fernandezianus .
Comparisons. Coryphaenoides leptolepis readily differs from other species of the subgenus Fuyangia , except C. brevibarbis , C. fernandezianus , and C. profundicola , in having the combination of 8–10 (usually 9 or 10) pelvic-fin rays and prominent pores along the cephalic sensory canals. Coryphaenoides leptolepis differs notably from the above three species in having a longer barbel (16–25% HL vs. <12%). It further differs from C. brevibarbis and C. fernandezianus in having a wider gill slit (22–28% HL vs. 15–19% in C. brevibarbis and 18% in C. fernandezianus ); from C. brevibarbis in having a narrower interorbital space (21–27% HL vs. 30–37%); from C. fernandezianus in having a narrower snout (internasal width 16–20% HL vs. 23%); and from C. profundicola in having a larger orbit (17–21% HL vs. 14%), smaller mouth (upper-jaw length 39–46% HL vs. 48%), and a shorter pectoral fin (51–65% HL vs. 73%).
Uncertain variants. A specimen collected from the south of Hokkaido (BSKU 96947, 80.9 mm HL, 384+ mm TL; Fig. 114 View FIGURE 114 ) differs from typical specimens of C. leptolepis in that the posterior ends of the occipital canals abut along the dorsal midline of the head, and there is no interspace between the occipital pores ( Fig. 113B View FIGURE 113 ). In contrast, these pores are always separated from each other in the other specimens of C. leptolepis examined ( Fig. 113A View FIGURE 113 ), with their interspace being 4–7% HL. Because such variation has not been reported for any species of grenadiers, this unusual specimen was initially considered to represent an undescribed species. However, similar variation was also found in the C. yaquinae examined (NSMT-P 78022, 82.7 mm HL, 441+ mm TL), and the difference in the occipital pores could be attributable to intraspecific variation. The Hokkaido specimen is further characterized by having a narrower gill slit (22% HL vs. 23–28% in the other C. leptolepis specimens examined), and fewer scale rows below the second dorsal-fin origin 7.5 (vs. 8–11.5) and above the anal-fin origin 22 (vs. 23– 34). However, these differences seem too subtle to warrant their specific separation, and the disjunct datasets may overlap if more specimens are included in the analysis.
Melo et al. (2010) reported Coryphaenoides sp. cf. leptolepis based on a small damaged specimen (MNRJ 26899, 302+ mm TL) collected from off Rio de Janeiro. This specimen differs notably from C. leptolepis in having lower counts of pelvic-fin rays [7 vs. usually 9–10 (ZMUB 3384 exceptionally has 8 rays on the left side)] and higher counts of first dorsal-fin rays [II,11 vs. II,8–10 (usually II,8–9; II,10 only in the paralectotype of C. liocephalus )]. Melo et al. (2010) also indicated that their specimen has a shorter pelvic fin as compared with published data for C. leptolepis (60% HL vs. 96%). This difference needs further confirmation, considering the poor condition of the additional Brazilian specimen and the fragile nature of the outer pelvic-fin ray. Its length also shows a positive allometry in C. leptolepis , with a wide range of variation (71–121% based on the present study). Due to the paucity of available materials, the taxonomic status of the additional Brazilian specimen is uncertain, but the differences in the fin-ray counts suggest it may be distinct from C. leptolepis . A definitive conclusion should be made when more specimens become available for comparisons.
Range extension. Coryphaenoides leptolepis was previously known only from the Atlantic and the North Pacific Oceans (see the synonymy of this species). During a visit to the Field Museum of Natural History (FMNH) in Chicago, a small nineteenth century specimen of C. leptolepis collected from the Indian Antarctic Ridge in the southeastern Indian Ocean (FMNH 5777, 39.3 mm HL, 137+ mm TL) was found among the collection. Surprisingly, the specimen was originally collected during the “ Challenger ” Expedition (sta. 158) in 1874. According to W. Leo Smith, a former curator of the FMNH fish collection, this historically important specimen was probably gifted to Chicago City for the World’s Columbian Exposition (also known as the Chicago World’s Fair) held in 1893. It represents not only the first record of C. leptolepis from the Indian Ocean, but also the second confirmed record of this species from the Southern Hemisphere. For future studies, counts and measurements for the Indian Ocean specimen are provided in Table 8 View TABLE 8 . The discovery of C. leptolepis from the Indian Ocean suggests that the species has a cosmopolitan distribution at bathyal depths of the world’s oceans.
In addition, three specimens collected from off Kesennuma, Miyagi Pref., at a depth of 7340–7433 m represent the deepest record for C. leptolepis , and confirm its occurrence in the hadal zone in the northwestern Pacific.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Coryphaenoides leptolepis Günther, 1877
| Nakayama, Naohide 2020 |
Coryphaenoides (Coryphaenoides) liocephalus
| : Okamura 1970 |
Coryphaenoides liocephalus:
| Gilbert & Hubbs 1916: 143 |
Coryphaenoides simulus:
| Gilbert & Hubbs 1916: 143 |
Coryphaenoides serrulus:
| Gilbert & Hubbs 1916: 144 |
Chalinura leptolepis:
| Goode & Bean 1896: 414 |
Chalinura serrula
| Bean 1890: 37 |
Chalinura simula
| Goode & Bean 1883: 199 |
Coryphaenoides leptolepis
| Gunther 1877 |
Co.leptolepis
| Gunther 1877 |
Ch. leptolepis
| Gunther 1877 |