
Amphisamytha vanuatuensis, Reuscher, Michael, Fiege, Dieter & Wehe, Thomas, 2009
|
publication ID |
https://doi.org/10.5281/zenodo.189379 |
|
DOI |
https://doi.org/10.5281/zenodo.6219214 |
|
persistent identifier |
https://treatment.plazi.org/id/B63287A9-FFD2-B74C-838E-FF39FCC7FABC |
|
treatment provided by |
Plazi |
|
scientific name |
Amphisamytha vanuatuensis |
| status |
sp. nov. |
Amphisamytha vanuatuensis View in CoL sp. nov.
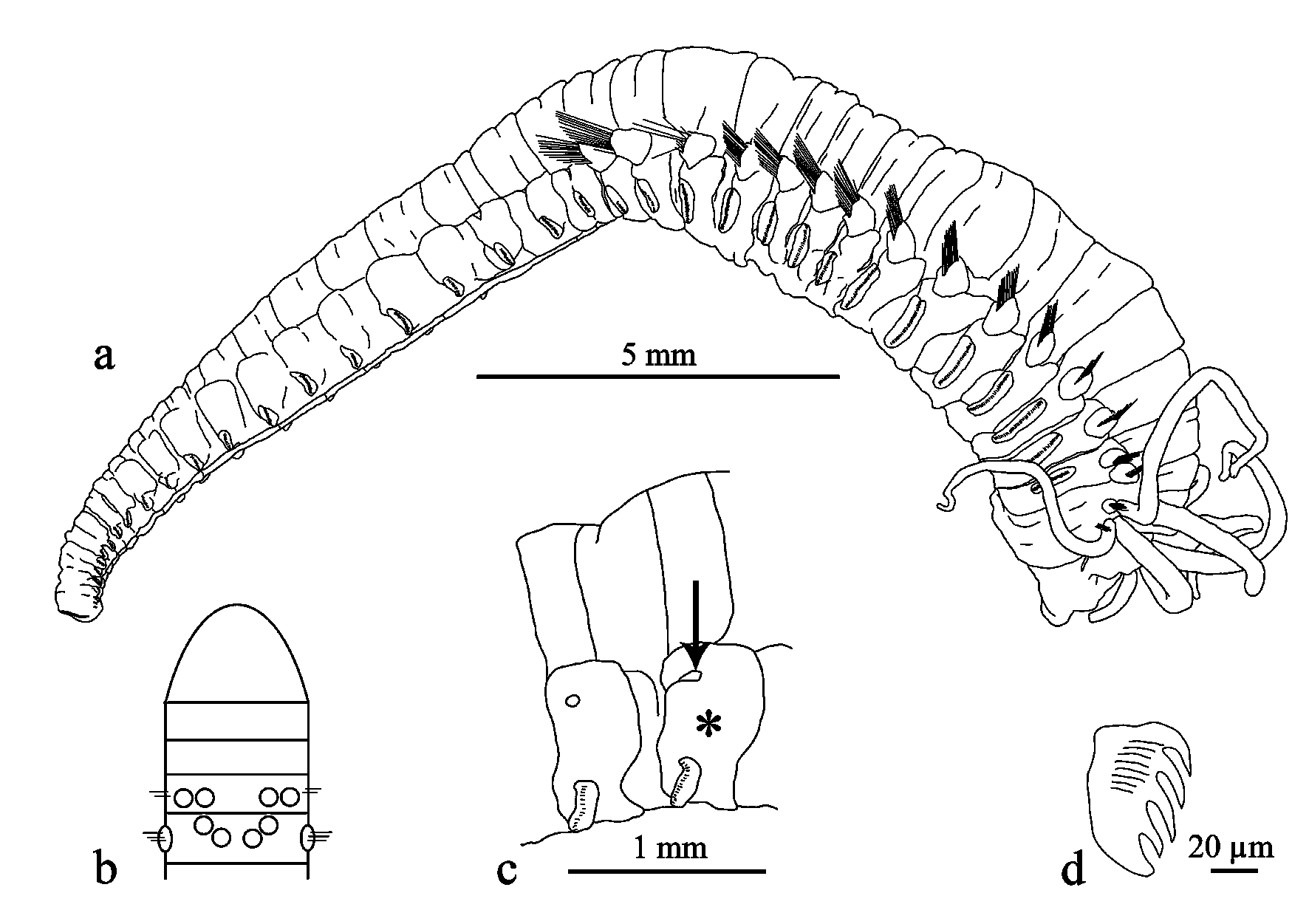
( Figs 2 View FIGURE 2 a–d, 3a–f; Tab. 4)
Specimens examined: Holotype, So 133, station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 1 cs (SMF 17798). Paratype, So 133, station 11 TVG, 3°19.058’S, 152°34.902’E, 1489 m, 22.7.1998, 1 cs (MNHN-Type 1496a). Paratypes, So 133, station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 2 cs (NHM ANEA 2009.18–19). Paratypes, So 133, station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 2 cs (SMF 17795). Paratypes, So 133, station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 2 cs (SMF 17794). Paratype, So 133, station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 1 cs (MNHN-Type 1496b). Paratypes, So 133, station 44 TVG, 3°19.352’S, 152°35.462’E, 1577 m, 26.7.1998, 2 cs (SMF 17799). Additional specimens: So 99: Station 91 TVG, 16°59.24’S, 173°55.00’E, 1983 m, 22.1.1995, 1 cs (SMF 17771). Station 98 TVG, 16°59.49’S, 173°54.83’E, 1992 m, 24.1.1995, 4 cs (SMF 17772). Station 115 TVG, 16°59.65’S, 173°54.73’E, 2003 m, 26.1.1995, 31 cs, 10 af (SMF 17769). So 133: Station 10 TVG, 3°18.855’S, 152°34.913’E, 1474 m, 22.7.1998, 1 af (SMF 17804). Station 11 TVG, 3°19.058’S, 152°34.902’E, 1489 m, 22.7.1998, 28 cs, 2 af (SMF 17807). Station 33 TVG, 3°19.097’S, 152°34.864’E, 1480 m, 25.7.1998, 170 cs, 77 af (SMF 17800). Station 34 TVG, 3°19.051’S, 152°34.864’E, 1481 m, 25.7.1998, 10 cs (SMF 17803). Station 35 TVG, 3°18.732’S, 152°39.491’E, 1114 m, 25.7.1998, 2 cs (SMF 17802). Station 44 TVG, 3°19.352’S, 152°35.462’E, 1577 m, 26.7.1998, 328 cs, 198 af (SMF 17808, SMF 17801 juveniles). So 134: Station 39 TVG, 16°59.348’S, 173°55.029’E, 1986 m, 21.8.1998, 3 cs (SMF 17776). Station 33 TVG, 16°59.454’S, 173°55.016’E, 1996 m, 20.8.1998, 16 cs, 3 af (SMF 17778). Station 35 TVG, 16°59.426’S, 173°54.819’E, 2002 m, 20.8.1998, 464 cs, 38 af (SMF 17774, SMF 17779 juveniles). Station 66 TVG, 16°59.447’S, 173°54.937’E, 1997 m, 25.8.1998, 239 cs, 14 af (SMF 17775, SMF 17781 juveniles). Station 99 TVG, 16°59.486’S, 173°54.910’E, 1999 m, 3.9.1998, 372 cs, 40 af (SMF 17777, SMF 17780 juveniles).
Additional type material examined: Amphisamytha fauchaldi Solís-Weiss & Hernández-Alcántara, 1994 . Holotype, Alvin Dive 1979, Guaymas Basin, Southern Trough, Riftia washings, 2014 m, 18.2.1988, 1 cs ( USNM 168087).
Amphisamytha galapagensis Zottoli, 1983 a. Holotype, Galapagos Rift Alvin Expedition, Alvin Dive N 990 #41, Galapagos Rift, Rose Garden, 2450 m, 7.12.1979, 1 cs ( USNM 81288).
Description. Length of holotype 20 mm; width (excl. parapodia) 3.5 mm. Body robust, epidermis leatherlike. Body from about unciniger 6 gradually tapering towards pygidium ( Fig. 2 View FIGURE 2 a). Prostomium trilobed, without eye-spots. Buccal tentacles withdrawn. Branchiae with deep furrows. First two pairs of branchiae in transverse row on chaetiger 1 (segment III), third pair of branchiae just posterior to inner branchiae of transverse row, fourth pair of branchiae medially and posterior to third. Third and fourth pair of branchiae arising from chaetiger 2 (segment IV) ( Fig. 2 View FIGURE 2 b). Thoracic chaetigers from segment III. Notopodia of first chaetiger reduced. Thoracic notopodia without cirri. Segments, especially abdominal ones, with transverse incision on dorsal side. One part of segment (thoracic and abdominal ones) cushion-like, glandular. Abdomen long, comprising about half of entire body length. 23 abdominal segments. Abdomen with ventral grove. Glandular pads above abdominal neuropodia, each bearing minute papilliform cirrus on dorsal side ( Fig. 2 View FIGURE 2 c). Anal cirri absent. Capillary notochaetae ( Fig. 3 View FIGURE 3 e) appearing bilimbate under light microscope. Thoracic and abdominal uncini similar in shape; four teeth in one vertical row above basal prow ( Fig. 2 View FIGURE 2 d).
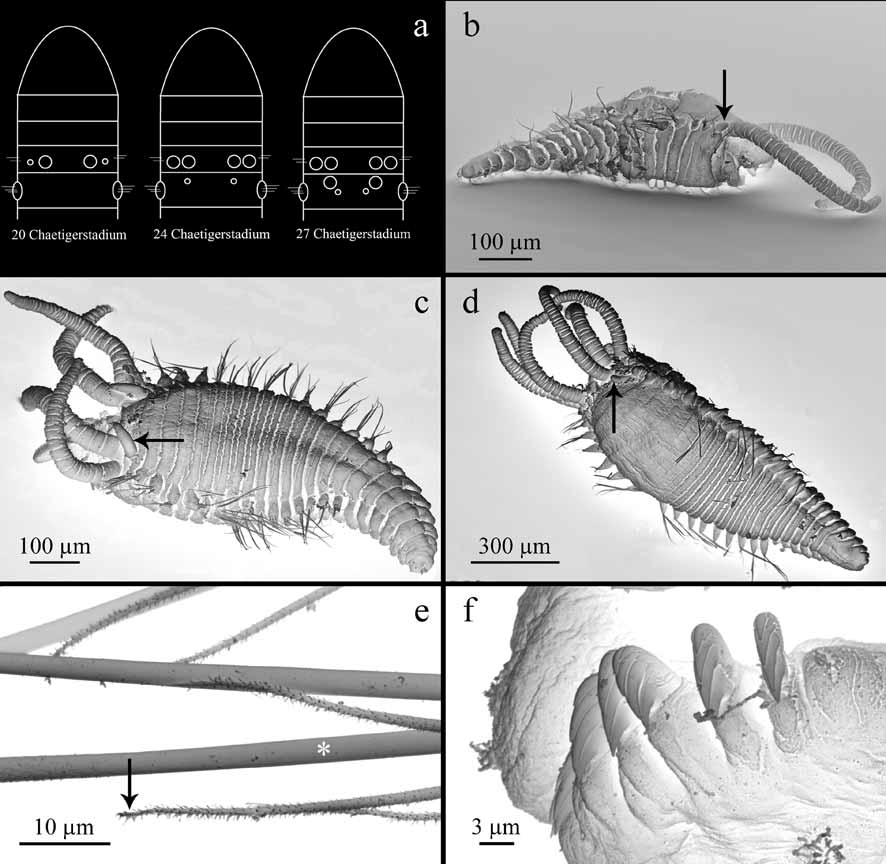
Variation. Range of body length 1–27 mm. Segment II of early juvenile stages (up to about 20 segment stage) with chaetae. The material studied here comprises many small and minute specimens with varying number of branchiae present: specimens with one to four pairs of branchiae at all stages of development ( Fig. 3 View FIGURE 3 a–d). The second pair of branchiae is visible from 20 chaetiger stage ( Fig. 3 View FIGURE 3 b), the third pair of branchiae from 24 chaetiger stage ( Fig. 3 View FIGURE 3 c), and fourth pair of branchiae from 27 chaetiger stage ( Fig. 3 View FIGURE 3 d).
Some specimens with buccal tentacles visible. They are smooth and bear a deep ventral groove.
Most common number of abdominal segments is 22, with the largest number found 25. Uncini of juvenile specimens have additional small lateral teeth ( Fig. 3 View FIGURE 3 f). These may be present along with the common type described above in one specimen and even in one torus whereas the uncini with the additional lateral teeth are always found at ventral part of the torus.
Remarks. This new species can easily be distinguished from A. galapagensis and A. fauchaldi by the longer abdomen and larger number of abdominal segments (Table 4). Amphisamytha vanuatuensis sp. nov., has up to 25 abdominal segments (the most common number is 22), while the maximum of abdominal segments in the two other species is 15. While even the largest specimens of A. galapagensis (ca. 10 mm) and A. fauchaldi (ca. 20 mm) never seem to exceed 15 abdominal segments, specimens of A. vanuatuensis sp. nov., have 15 segments measuring at maximum 5 mm. Specimens of 10 mm already have 20 or more abdominal segments. Furthermore, Zottoli (1983a) described for A. galapagensis the appearance of the second pair of branchiae in the 14 chaetiger stage, while we observed it in the 20 chaetiger stage for A. vanuatuensis sp. nov., ( Fig. 3 View FIGURE 3 b). Fortunately, we had the opportunity of studying a large amount of juvenile and adult specimens (> 1900 specimens) and could therefore examine the development of branchiae in detail and correlate their first appearance to the respective chaetiger stages ( Fig. 3 View FIGURE 3 a–d).
It appears that the species of Amphisamytha occurring at hot vents might have evolved by allopatric speciation, because A. vanuatuensis sp. nov., was found exclusively in the Western Pacific. Records of A. galapagensis from the Western Pacific ought to be rechecked, since they may represent specimens of A. vanuatuensis sp. nov. It appears that A. galapagensis is neither a cosmopolitan, as suggested by Petrecca & Grassle (1987) and Galkin (1997), nor a pan–Pacific species, as suggested by McHugh & Tunnicliffe (1994), but rather confined to the Eastern Pacific in its distributional range.
Type material of Amphisamytha fauchaldi and A. galapagensis was examined to assess the justification of the description of a new species and the validity of of A. fauchaldi . A. vanuatuensis sp. nov., is considered a new species as discussed above, A. fauchaldi is considered here to be a junior synonym of A. galapagensis . A. fauchaldi has been described exclusively from the hot vents of Guaymas Basin, a location right in the middle of the distribution area of A. galapagensis . The characters used to distinguish the two species, i.e. a smaller gap between the groups of branchiae in A. fauchaldi , relative branchial length to body length, 3: 4 in A. fauchaldi and 1: 3 in A. galapagensis , and larger body size of A. fauchaldi , appear to be inadequate since they vary with preservation, development and age. The larger specimens described as A. fauchaldi are likely to be older specimens of A. galapagensis . This reasoning is supported by the examination of the holotypes of both Eastern Pacific species.
Doubts about the wide distribution of A. galapagensis in the Pacific were noted by Blake (1991), when he described specimens from the Mariana Basin with an unusual number of 17–18 abdominal segments. Blake explained this phenomenon and the additional teeth of the specimens' uncini as genetic differences between populations. The uncini's shape, however, is typical for juvenile specimens adding weight to the assumption that the specimens described by Blake, are juveniles and therefore the full complement of abdominal segments had not been attained. Thus, we consider these specimens as belonging to Amphisamytha vanuatuensis sp. nov., rather than A. galapagensis .
Two juvenile specimens from LHOS Area A (North Fiji Basin) with one and two attached parasitic copepods, respectively were found in the sample. This phenomenon is well-known from hot vent ecosystems ( Heptner & Ivanenko 2002, Desbruyères et al. 2006).
Etymology. The species name refers to the insular state Vanuatu near the sampling site in the North Fiji Basin. The name emphasizes the Western Pacific distribution of this species in contrast to its Eastern Pacific sister-species A. galapagensis .
Distribution. West Pacific: Lihir Basin, North Fiji Basin.
Table 4: Distinguishing characters of the four Amphisamytha species considered valid (Data for A. bioculata taken from Williams (1987) and Hilbig (2000), for A. japonica from original description, and for A. galapagensis from the original description supplemented by own observations)
Relation of ?? 1:3 - 3:4 1:2 - 1:4 branchial length to
body length
Appearance of 2nd ?? 14 chaetiger stage 20 chaetiger stage pair of branchiae
Abdominal 13-14 14 12-15 up to 25 segments
Number of ?? 10-13 10-15 abdominal segments
of specimens <5 mm
Number of ?? 15 21-25 abdominal segments
of specimens> 10
mm
Specialized 1st pair inconspicuous 1st and last two pairs 1st pair inconspicuous 1st pair inconspicuous notopodia inconspicuous
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ampharetinae |
|
Genus |