
Mustela africana Desmarest, 1818
|
publication ID |
https://doi.org/10.1644/917.1 |
|
persistent identifier |
https://treatment.plazi.org/id/B60087D1-4A5F-FFE4-9B9C-FC3AFC94FDC7 |
|
treatment provided by |
Carolina |
|
scientific name |
Mustela africana Desmarest, 1818 |
| status |
|
Mustela africana Desmarest, 1818 View in CoL
Amazon Weasel
Mustela africana Desmarest, 1818:376 View in CoL . Type locality ‘‘Afrique,’’ corrected to ‘‘los arrabales de Belem, la antigua Para´, [ Brazil],’’ by Cabrera (1958:257).
Gymnopus africanus: Gray, 1865:120 . Name combination.
Mustela stolzmanni Taczanowski, 1881:835 View in CoL . Type locality ‘‘Peruvia orient. Yurimaguas.’’
Putorius brasiliensis paraensis Goeldi, 1898:560 . Type locality ‘‘Stadt Para´ (Waldrevier von Marco da Legoa),’’ [ Brazil]; corrected to ‘‘matas do rio Murutucu, arrabalde de Belém, Para´,’’ by Carvalho (1959:461).
[ Putorius View in CoL ] [ brasiliensis ] paraënsis Goeldi, 1898:562 . Unjustified emendation of Putorius brasiliensis paraensis Goeldi, 1898 .
Putorius ( Mustela) brasiliensis paräensis: Goeldi, 1898:562 . Name combination.
Putorius paraensis: Goeldi and Hagmann, 1904:61 . Name combination.
Mustela paraensis: Cabrera, 1913:432 . Name combination.
[ Mustela View in CoL ] pacaensis Cabrera, 1914:176. Incorrect subsequent spelling of Mustela paraensis Goeldi, 1898 .
Mustela affinis paraensis: Hollister, 1914:143 . Name combination.
Mustela stolzmanni paraensis: Hall, 1936:111 . Name combination.
Mustela stolzmanni stolzmanni: Hall, 1936:111 . Name combination.
Mustela africana africana: Hall, 1939:159 View in CoL . First use of current name combination.
Mustela africana stolzmanni: Hall, 1939:159 View in CoL . Name combination.
[ Grammogale View in CoL ] africana: Cabrera, 1940:15 View in CoL . Name combination.
Grammogale africana: Cabrera and Yepes, 1940:150 . Name combination.
Mustela paraense Cabrera and Yepes, 1940:151 . Incorrect subsequent spelling of Mustela paraensis Goeldi, 1898 .
Grammogale africana africana: Cabrera, 1958:257 . Name combination.
Grammogale africana stolzmanni: Cabrera, 1958:257 . Name combination.
Mustela ( Grammogale) africana africana: Pine, 1973:71 View in CoL . Name combination.
Mustela ( Grammogale) africana: Izor and Peterson, 1985:788 View in CoL . Name combination.
CONTEXT AND CONTENT. Order Carnivora View in CoL , suborder Caniformia View in CoL , family Mustelidae View in CoL , subfamily Mustelinae View in CoL , genus Mustela View in CoL . Two subspecies are recognized ( Wozencraft 2005):
M. a. africana Desmarest, 1818:376 . See above.
M. a. stolzmanni Taczanowski, 1881:835 . See above.
NOMENCLATURAL NOTES. Mustela africana has been includ- ed in the subgenus Grammogale Cabrera, 1940 , together with M. felipei (Izor and de la Torre 1978; Youngman 1982), and the case for recognizing Grammogale as a distinct genus has been discussed (Cabrera 1940; Izor and de la Torre 1978; Izor and Peterson 1985). Bacular characteristics separate the species of Grammogale from all other extant New World mustelids (Izor and Peterson 1985). Baryshnikov and Abramov (1997) and Abramov (2000) considered Grammogale a monotypic subgenus containing only M. africana , and proposed the subgenus Cabreragale for M. felipei and the genus Neovison for the American mink. Their proposal for mink has been generally accepted (e.g., Wozencraft 2005; Larivière and Jennings 2009). In a phylogenetic analysis of the genus Mustela based on cytochrome- b sequences, Harding and Smith (2009) recovered a grouping of M. africana , M. felipei , M. frenata , and Neovison vison . If confirmed by additional analyses, this set of relationships would restrict the genus Mustela to weasels and stoats with Eurasian–Holarctic distributions, and Grammogale would replace Neovison as the subgeneric or generic name for this set of mustelids.
The specific epithet name, africana , was given by mistake because the specimen, taken to Paris in an exchange from the ‘‘Cabinet de Lisbonne,’’ Museum of Ajuda in Portugal, was thought to originate from Africa. However, in 1913 it was proven that it pertained instead to the tropical weasel of Brazil ( Cabrera 1913, 1914; Hall 1951). The type locality was assumed to be Para´( Hall 1951), and later fixed as ‘‘los arrabales de Belem, la antigua Para´,’’ Brazil ( Cabrera 1958). English common names are Amazon weasel and tropical weasel; local names include doninha-amazônica in Brazil ( Paglia et al. 2012); aitaka chukuri, chucuri grande, gombe, kujamchana, maña yai, and totosi in Ecuador ( Tirira 2004, 2007); and comadreja amazónica, comadreja rayada, and katori in Peru ( Pacheco et al. 2009).
DIAGNOSIS
Body size of Mustela africana averages larger than that of the other South American weasels, M. felipei (Colombian weasel) and M. frenata (long-tailed weasel— Hall 1951; Izor and de la Torre 1978), reaching about 500 mm in total length, versus 350 mm and 420 mm, respectively. M. africana exhibits a ventral stripe that is the same color as the dorsum ( Fig. 1 View Fig ). M. felipei has a similar ventral marking but it is reduced to a spot on the chest or neck (Ramírez- Chaves et al. 2012) and M. frenata has no ventral markings. The tail is fairly long for a weasel (Ž 50% head-and-body length) and uniform in color. The soles of the feet lack fur and a thenar pad is present on forefoot ( Hall 1951). The skull of M. africana ( Fig. 2 View Fig ) has a mesopterygoid fossa reduced in comparison with M. felipei , and the auditory bullae are narrow, widely spaced, elongated, and less inflated than in M. frenata ( Hall 1951; Izor and de la Torre 1978; Abramow 2000). The nasals form an isosceles triangle, in contrast with M. felipei and M. frenata in which the lateral margins are subparallel anteriorly. The p2 is absent in M. africana (Izor and de la Torre 1978) .
GENERAL CHARACTERS
Mustela africana presents an elongate body and neck, small head, and short ears and legs. The pelage is short, uniformly chestnut brown or reddish with little variation along the body but the nose is darker. As in other Neotropical weasels, young individuals have darker pelage than adults, and pelage color varies geographically, with western populations dark brown and eastern populations more reddish. The venter is pale orange-tan with a midventral stripe of the same color as the dorsum ( Taczanowski 1881; Pine 1973). The ventral stripe can extend to the forelegs or the throat and in some specimens it extends to the base of the chest and then splits into 2 or 3 semicontinuous segments ending on the neck ( Orcés 1944; Izor and de la Torre 1978). The facial vibrissae are short and do not reach the posterior margin of the ear ( Taczanowski 1881; Hall 1951). The soles of the feet are naked, with a few scattered hairs on ventral sides of interdigital membranes ( Hall 1951). external measurements (mm; with parenthetical ranges [when appropriate], n —compiled from Taczanowski 1881; Goeldi 1901; Goeldi and Hagmann 1904; Hall 1951; Izor and de la Torre 1978; Izor and Peterson 1985) of adult (mixed sexes) of M. a. stolzmanni , were: total length, 476 (430–523, 2), 425 (415–435, 2); length of tail, 175 (160–190, 2); length of hind foot, 53 (52–54, 3); length of ear, 21 (20– 22, 2). Additional external measurements are available ( Taczanowski 1881; Goeldi 1901; Goeldi and Hagmann 1904; Hall 1951; Izor and de la Torre 1978; Izor and Peterson 1985).
The skull presents a large and rounded braincase, anteriorly inflated; wide palatal region; tympanic bullae less inflated than other South American species of the genus; and a reduced angle of lower jaw. The dentary is heavy with the masseteric fossa not extending anteriorly to the posterior one-fourth of talonid of ml ( Hall 1951). Mean craniodental measurements (mm; with parenthetical range—compiled from Goeldi and Hagmann 1904; Hall 1951; Izor and Peterson 1985) of 3 males of M. a. africana were: condylobasal length, 45.8 (44.6–47.8); zygomatic breadth, 29.5 (26.8–32.2); least interorbital breadth, 11.3 (10.2–12.9); length of upper toothrow, 16.7 (16.4–17.2). For adult males and females of M. a. stoltzmanni mean craniodental measurements (mm, parenthetical ranges [when appropriate], n) were: condylobasal length, 51.2 ( 1 male), 46.7 (44.6– 49.8, 3 females); zygomatic breadth, 30.4 ( 1 male), 28.86 (28.81–28.90, 2 females); least interorbital breadth, 12.2 ( 1 male), 11.3 (11.0–12.0, 3 females); length of upper toothrow, 16.1 ( 1 male), 16.8 (16.0–17.5, 3 females). Additional cranial measurements are available (Goeldi and Hagmann 1904; Hall 1951; Izor and Peterson 1985).
Mean external measurements (mm; with parenthetical ranges [when appropriate], n —compiled from Taczanowski 1881; Goeldi 1901; Goeldi and Hagmann 1904; Hall 1951; Izor and de la Torre 1978; Izor and Peterson 1985) of adult (mixed sexes) M. a. africana were: total length, 510 (430– 522, 7); length of tail, 200 (2), 192 (168–210, 4); length of hind foot, 53.3 (50–56, 3); length of ear, 18 (2). Mean
DISTRIBUTION
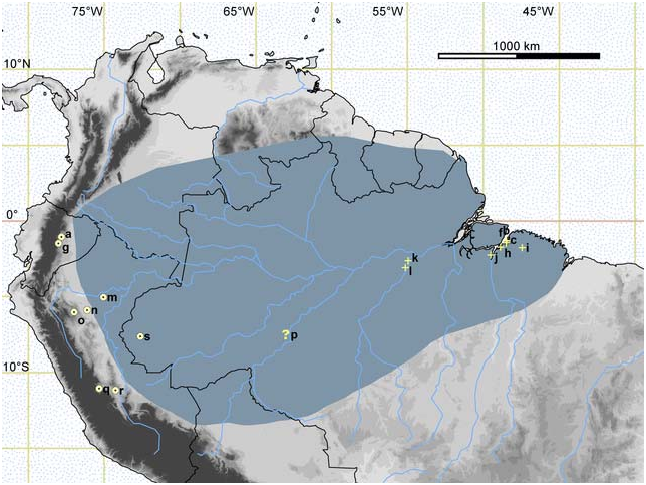
Mustela africana is known from few localities ( Fig. 3 View Fig ) located south of the Amazon River in Brazil and its tributaries in eastern Ecuador and Peru (the Napo, the Marañón, and the Ucayali) over an elevational range from 0 to 1,250 m ( Schreiber et al. 1989; Tirira 2007; Ramírez- Chaves et al. 2012). The limits of the range of distribution are poorly known and the species may be distributed in the Amazon Basin of Colombia and northern Bolivia ( Alberico et al. 2000; Emmons and Helgen 2008). No fossils are known.
FORM AND FUNCTION
The baculum of Mustela africana has a slender shaft that becomes broad and shallow in its distal part and possesses wide margins at the level of the dorsal bend. It presents a trifid tip with the medial and right lateral processes connected by a thin and translucent layer of bone. The glenoid fossa (Izor and de la Torre 1978). A female from Moyobamba, Peru, deposited at the Natural History Museum (BMNH 24.12.12.24) has 3 pairs of mammae, 1 pair inguinal and the other 2 on the posterior part of the abdomen ( Hall 1951).
trifid tip is a character shared with M. felipei (Izor and Peterson 1985) .
The dental formula is i 3/3, c 1/1, p 2–3/2, m 1/2, total 30–32, and the dentition is specialized for a carnivorous diet. The teeth are robust; the medial lobe of Ml is slightly larger than the lateral lobe; and the deuterocone of P4 is heavy and usually inclined mesially ( Hall 1951). P2 can be absent or present, and is generally absent in M. a. stolzmanni (Izor and de la Torre 1978) . The p2 is absent; a small p2 is present in Colombian specimens of both M. felipei and M. frenata .
In mature M. africana , the 2 temporalis muscles meet at the midline of the skull at or anterior to the level of the
ECOLOGY
Little is known of the habitat preferences and natural history of Mustela africana , apart from the places where the specimens have been found. Semiaquatic life has been suggested by its naked foot soles with interdigital webbing ( Schreiber et al. 1989), although these traits are found in other mustelids (e.g., Gulo and Martes ) with terrestrial and arboreal habits. More direct evidence comes from observations of its swimming abilities; a specimen in the Goeldi Museum was captured during the day while swimming in a brackish estuary in Pará, Brazil, nearly one-half mile from shore ( Tate 1931). Four weasels reported as this species were sighted foraging together in the lower canopy of forest along the Rio Madeira (7833 0 S, 63812 0 W), Amazonas, Brazil, but the identification of these Mustela could not be confirmed (Ferrari and Lopes 1992). Specimens also were found in a willow tree in Moyobamba, Peru ( Hall 1951), in a rubber plantation in Brazil (Izor and Peterson 1985), and burrowing in a hollow tree stump (Emmons and Feer 1990). One individual was found in humid forest of Maynas, Loreto, Peru, at an elevation of 150 m ( Taczanowski 1881; Hall 1951). A specimen was captured alive in a trap baited with maize and banana and set near a house, in a rubber plantation located in Belterra, Pará, Brazil (Izor and Peterson 1985).
GENETICS
Phylogenetic analysis using sequences of the mitochondrial cytochrome- b indicates that Mustela africana is the sister species of M. felipei , with a divergence time from M. frenata estimated at about 3–4 million years ago. These 3 species, together with Neovison vison (formerly Mustela ), comprise a distinct New World lineage that is sister to 2 Southeastern Asian species and then to a large diverse Holarctic lineage (Harding and Smith 2009). The cytochrome- b sequence of M. africana was obtained from a Brazilian specimen (FMNH 106488) housed at the Field Museum of Natural History, Chicago, Illinois.
CONSERVATION
Mustela africana is only known from a few specimens but seemingly is widespread throughout Amazonia (Schreib- er et al. 1989; Oliveira 2009). Globally, it is considered ‘‘Least Concern’’ by the International Union for Conservation of Nature and Natural Resources; however, it may qualify as ‘‘Data Deficient’’ because almost nothing is known of its ecology, threats, and distribution (Emmons and Helgen 2008). Previous assessments for Brazil and Ecuador included the species in ‘‘Data Deficient’’ based on the unknown status and apparent rarity of the species ( Machado et al. 2005; Tirira 2007). The species was considered the least known carnivore of Brazil ( Oliveira 2009), although it may occur in large national parks of Amazonia ( Schreiber et al. 1989), including National Parks Sumaco-Napo Galeras and Yasuní in Ecuador ( Tirira 2007).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mustela africana Desmarest, 1818
| Ramírez-Chaves, Héctor E., Arango-Guerra, Heidi Liliana & Patterson, Bruce D. 2014 |
Mustela ( Grammogale ) africana africana :
| PINE, R 1973: 71 |
Grammogale africana africana :
| CABRERA, A 1958: 257 |
Grammogale africana stolzmanni :
| CABRERA, A 1958: 257 |
Mustela africana africana :
| HALL, E 1939: 159 |
Mustela africana stolzmanni :
| HALL, E 1939: 159 |
Mustela stolzmanni paraensis :
| HALL, E 1936: 111 |
Mustela stolzmanni stolzmanni :
| HALL, E 1936: 111 |
Mustela
| CABRERA, A 1914: 176 |
Mustela affinis paraensis :
| HOLLISTER, H 1914: 143 |
Mustela paraensis :
| CABRERA, A 1913: 432 |
Putorius brasiliensis paraensis Goeldi, 1898:560
| CARVALHO, C 1959: 461 |
| GOELDI, E 1898: 560 |
Putorius
| GOELDI, E 1898: 562 |
Putorius ( Mustela ) brasiliensis paräensis : Goeldi, 1898:562
| GOELDI, E 1898: 562 |
Mustela stolzmanni
| TACZANOWSKI, L 1881: 835 |
Gymnopus africanus :
| GRAY, J 1865: 120 |
Mustela africana
| CABRERA, A 1958: 257 |
| DESMAREST, A 1818: 376 |