
Glaucalges tytonis, Dabert, Jacek, Ehrnsberger, Rainer & Dabert, Miroslawa, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181058 |
|
DOI |
https://doi.org/10.5281/zenodo.6227053 |
|
persistent identifier |
https://treatment.plazi.org/id/B4608780-FFF3-FFB9-FF1D-FC9494E1F809 |
|
treatment provided by |
Plazi |
|
scientific name |
Glaucalges tytonis |
| status |
sp. nov. |
Glaucalges tytonis sp. nov.
Material examined. Type material: 1 male holotype, 3 male, 3 female paratypes from Tyto alba ( Strigiformes , Tytonidae ), near Osnabrueck (52.16 N 0 8.03 E), Germany, 12 November 1992, no other data; DNA barcode GenBank Acc. EU271955 View Materials for male holotype and 3 male paratypes, voucher AMUFM 636; remaining paratypes vouchers AMUFM 637–639.
Additional material: 17 males from same host (another bird individual), same data; DNA barcode Gen- Bank Acc. EU271956 View Materials for 13 males, voucher AMUFM 795; remaining individuals vouchers AMUFM 791– 794.
Comparative material: Glaucalges attenuatus , two samples: (1) 9 males from Asio otus (type host species), near Osnabrueck, Germany, 26 February 1993, no other data; DNA barcode GenBank Acc. EU271958 View Materials for 8 males, voucher AMUFM 780, (2) 3 males, 2 females from same host, Poznan (52.25 N 16.52 E), Poland, 11 May 2005, coll. A. Badek; DNA barcode GenBank Acc. EU271957 View Materials for all individuals, voucher AMUFM 211.
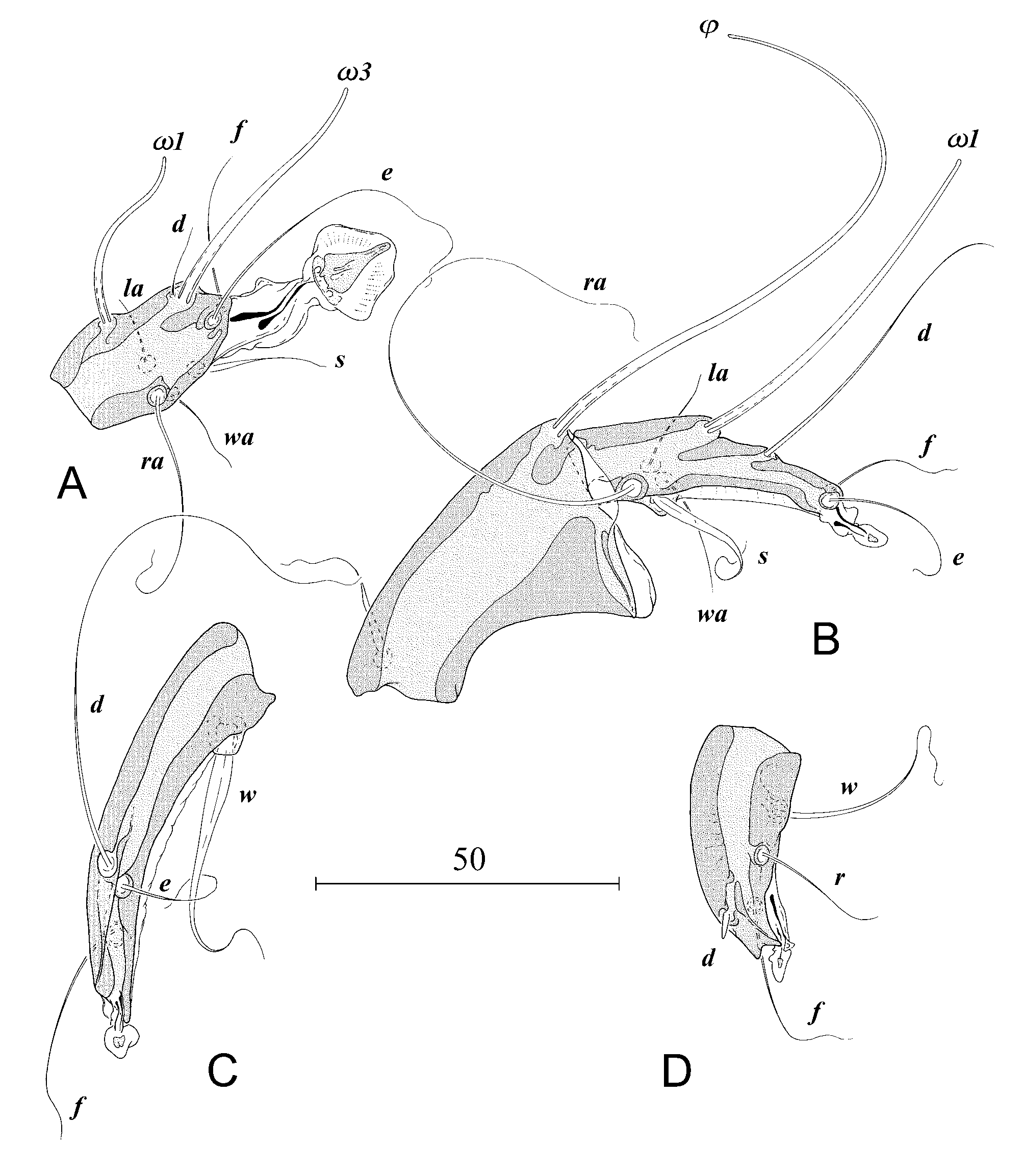
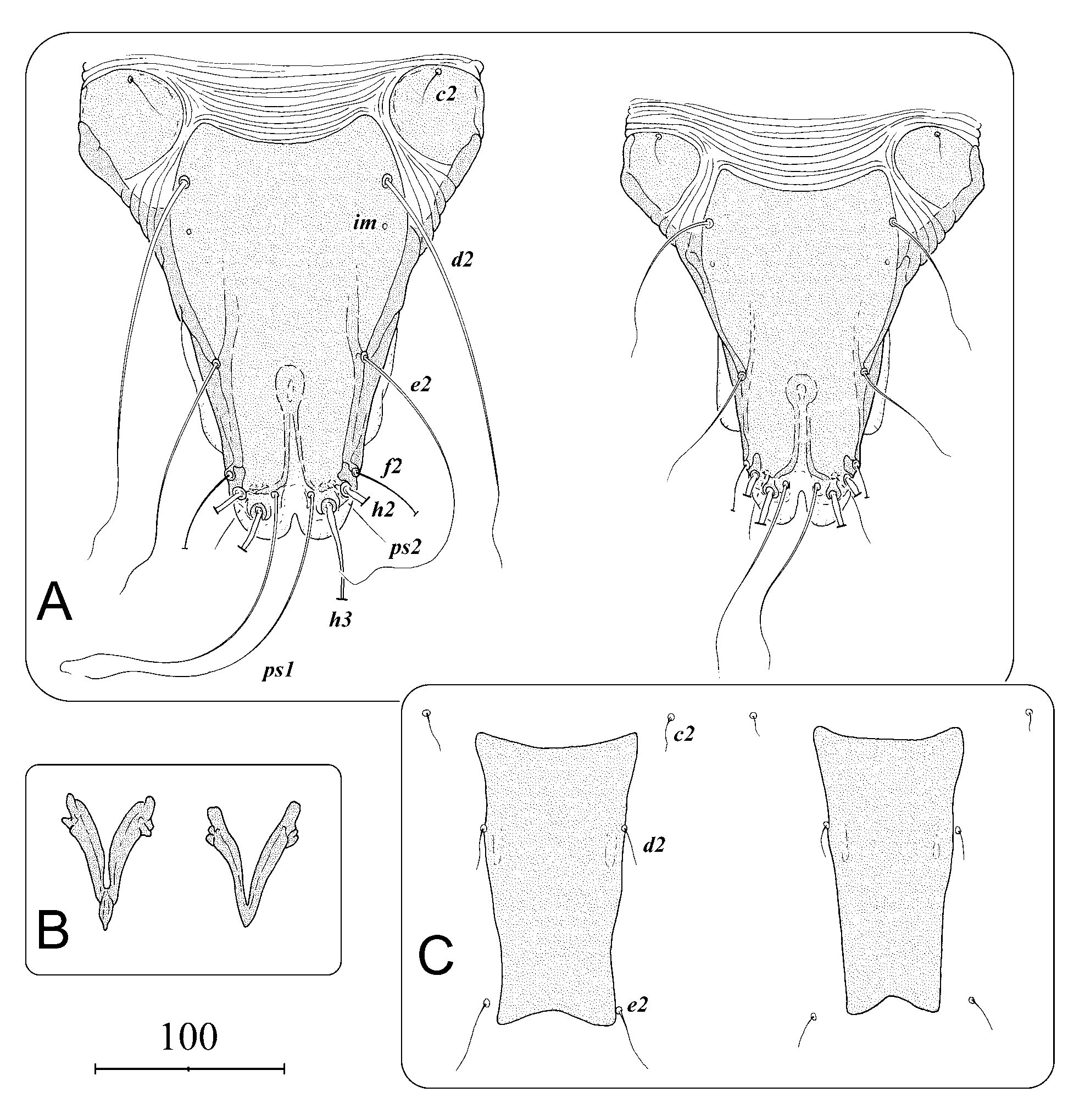
Description. Male ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 5 View FIGURE 5 A–B, 6, 7). Gnathosoma with rectangular subcapitulum and finger-like appendages on lateral margins, length of gnathosoma 50 (48–54), width 48 (46–50), length/width 1.0 (1.0– 1.1). Idiosoma relatively stout, strongly narrowing terminally and shaped as inverted triangle, length 368 (320–375), width 243 (174–255), length/width 1.5 (1.5–1.8). Propodosoma and hysterosoma lengths 110 (96– 115) and 258 (224–260) respectively, propodosoma/hysterosoma 0.4. Terminal margin of opisthosoma with elongated trapezoidal opisthosomal lobes fused medially. Remnants of interlobar cleft are connected with posterior part of ovate supranal concavity in a keyhole-like structure. Length of the cleft including supranal concavity 80 (62–87), width at the level of setae h2 bases 11 (9–14). Terminal margins of lobes with tongue-like rounded membranes separated by roundly triangular incision reaching anteriorly level of insertion of setae h3. Lateral opisthosoma with narrow membranes extending from bases of legs IV to level anterior to bases of setae f2. Pronotal shield triangular in shape anteriorly to level of setae si, posteriorly to this level rectangular; length of the shield 105 (97–112), maximal width 90 (82–90). Shield uniformly dotted, occupying medial part of dorsal propodosoma. Scapular shields well developed, shaped as quadrangles. Setae se shaped as macrosetae, set on striated tegument between humeral and pronotal shields. Setae si minute, set on lateral margins of pronotal shield. Humeral shields large, rounded with medium-sized setae c2 at anterior margins. Hysteronotal shield well developed and covers most of dorsal surface of hysterosoma; anterior margin concave. Maximal width of the shield 115 (109–122). The shield uniformly dotted with two longitudinal furrows extending from the level posterior to cupules im to bases of setae e2. Dorsal setae c1, d1, e1, h1 absent. Setae ps1 longer than distance d2–d2. Lateral setae d2, e2, f2 long; d2 and e2 extending beyond the level of tips of terminal membranes ( Fig. 4 View FIGURE 4 A). Setae h2 and h3 shaped as macrosetae longer than idiosoma. All dorsal and lateral setae piliform. Openings of opisthonotal glands gl absent. Only cupules im visible, set posterior to bases of setae d2. Distances between setae: c2–c2 171 (136–186), c2–d2 62 (51–68), d2–d2 107 (100–108), d2– e2 90 (89–102), e2– e2 78 (71–77), h3–h3 37 (35–39), ps1–ps1 20 (18–19).
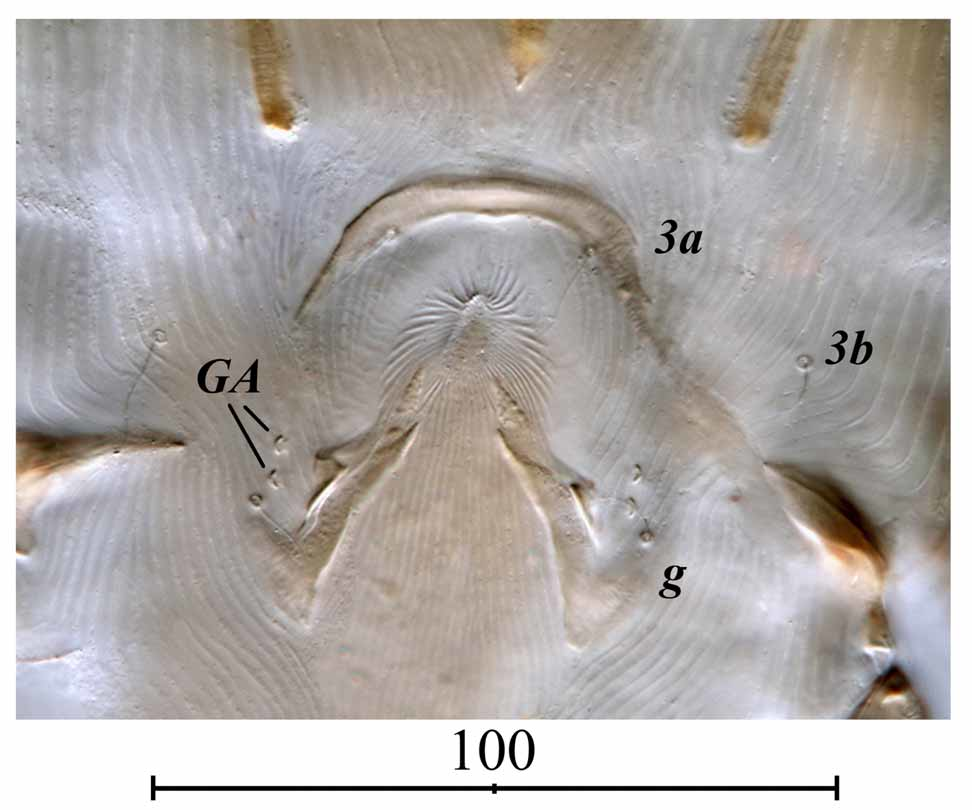
Epimerites I fused as Y with very short sternum ( Fig. 5 View FIGURE 5 B). Coxal fields of ventral propodosoma and coxal fields III without sclerotized shields near epimerites. Coxal fields IV completely covered by well sclerotized shields with medial expansions encompassing bases of setae 3a; in more weakly sclerotized individuals these expansions may be partly or completely reduced. Genito-anal region without cuticular shields with exception of minute square sclerite between setae ps3. Genital arch flat, length 11 (10–12), width 16 (14–15) with very short aedeagus. Genital acetabula set on epimerites IV laterally to genital organ. Setae 3b shaped as long macrosetae reaching by tips bases of f2. Setae ps2 at least as long as distance between them. Adanal discs slightly elliptical, diameter 13x12 (12x11–15 x12); each disc with small antero-medial sclerite. Corollae of discs radially striated. Distances between setae: 3b–3b 120 (64–111), 3b–3a 41 (30–44), 3a–g 68 (50–66), g–ps3 38 (35–42), ps3–ps2 70 (68–74).
Legs I–II of similar size, legs III longer than I–II, legs IV thicker than others. Tibiae I and II with ventral triangular apophyses. Femoragenua I and II with rounded dorsal retrograde apophyses. Tarsi II and III elongated with narrow ventral lamella. Tarsus IV short with two apical spines, tarsus III with single apical spine. Ambulacra I well developed, II–IV vestigial. Tips of leg IV tibiae reaching level of posterior margins of terminal membranes. Setae ba absent on tarsi I–II, r, s on tarsi III, and r, e on tarsi IV; setae d on tarsi IV sticklike, very short ( Fig. 2 View FIGURE 2 A–D).
Female ( Figs 3 View FIGURE 3 , 4 View FIGURE 4 , 5 View FIGURE 5 C). Gnathosoma shaped as in males, length 48–50, width 47–50, length/width 1.0– 1.1. Idiosoma moderately elongated, parallel-sided, length 310–315, width 149–160, length/width 2.0–2.1.
Propodosoma and hysterosoma lengths 92–105 and 210–218 respectively, propodosoma/hysterosoma 0.4– 0.5. Rounded or concave terminal margin with small terminal protuberance with copulatory opening on its tip; in some individuals copulatory opening situated more dorsally in which case protuberance is absent. Pronotum shaped and sclerotized as in male, pronotal shield length 92–96, width 85–92. Humeral shields absent. Hysteronotal shield shaped as strongly elongated trapezium, anterior margin longer than posterior one, both margins concave. Shield extends posteriorly slightly beyond the level of setae e2; length of the shield 149– 153, width at anterior margin 78–83. Two elongated lacunae lying medially to bases of setae d2. All dorsal shields uniformly dotted. Setae of pronotum as in males. Setae of dorsal hysterosoma short and with exception of d2 inserted on striated tegument outside the hysteronotal shield; setae d 2 set on lateral margins of the shield. Setae e1 nearly twice as long as d2 ( Fig. 5 View FIGURE 5 C). Distances between setae: c2–c2 125–135, c2–d2 64–68, d2–d2 70–74, d2– e2 91–92, e2– e2 67–76, h3–h3 44–57.
Ventral propodosoma shaped and sclerotized as in male. Epigynum well developed, arched, width 42–43, height 18–22 (max. 30 in additional material) ( Figs 3 View FIGURE 3 A, 4). Branches of epigynum encompassing bases of setae 3a. Latigenital sclerites extending posteriorly beyond the level of genital setae g. Setae g inserted posterior to the level of genital acetabula and posterior to the level of setae 3b. Setae 3b anterior to genital acetabula. Setae ps2 inserted latero-terminal to ps3, ps3 subequal to ps 2 in length.
Ambulacra of legs IV reaching terminal end of the body, legs III do not reach body terminus. Both posterior tarsi attenuate with narrow ventral lamella.
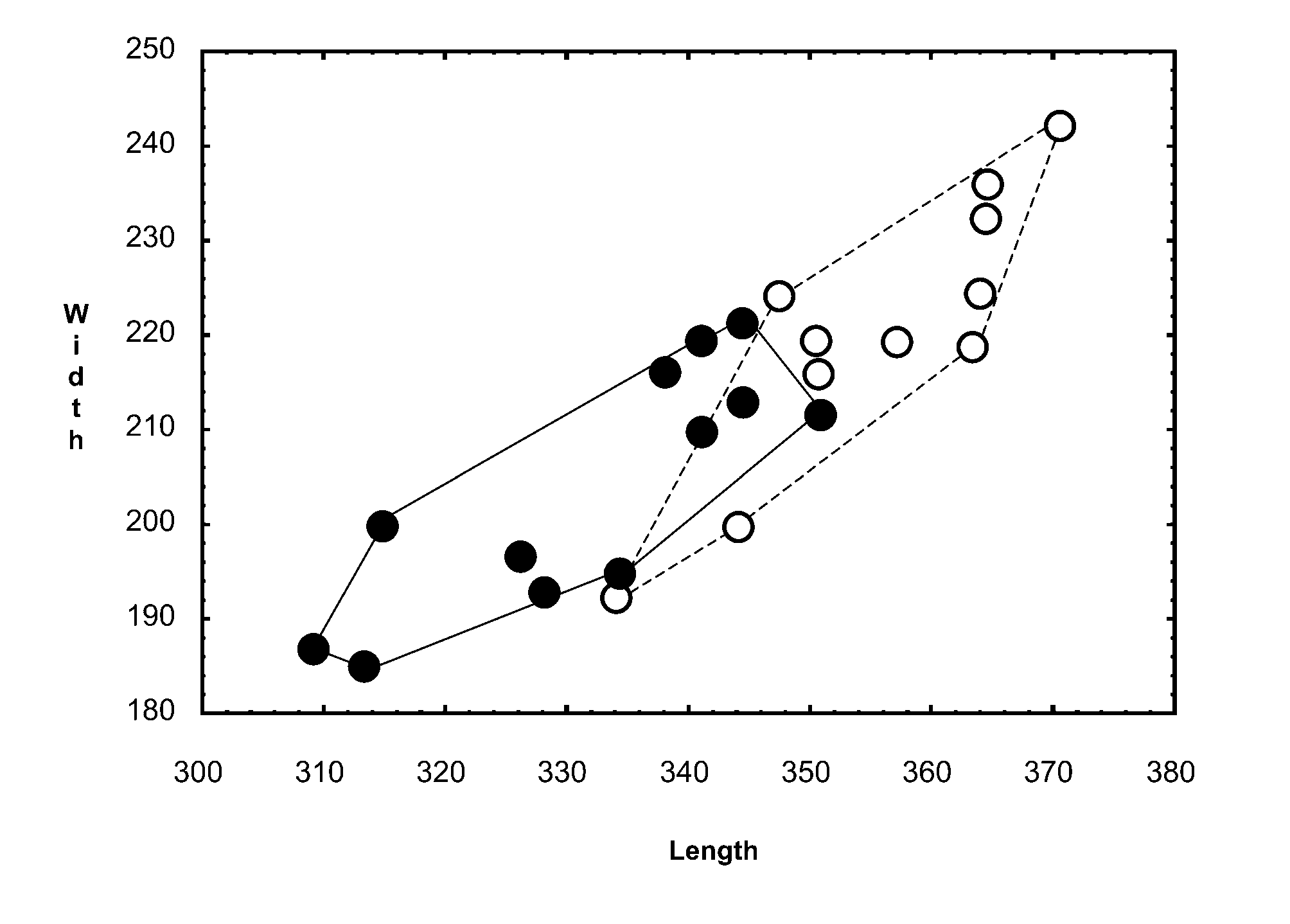
Differential diagnosis. Morphology. The new species is extremely similar to Glaucalges attenuatus ( Buchholz, 1869) described from the long-eared owl Asio otus ( Strigiformes , Strigidae ). Males of both species differ by length of dorsal setae d2 and e2: in G. tytonis sp. n. these setae are much longer than in G. attenuatus ( Figs 5 View FIGURE 5 A, 6). Females are almost indistinguishable. The only difference is in the relative lengths of setae d2 and e2: in females of G. t y t o n i s sp. n. setae e2 are clearly almost twice as long as setae d2; in G. attenuatus both setae are subequal ( Fig. 5 View FIGURE 5 C). Both sexes of G. tytonis sp. n. have Y-shaped epimerites I with short sternum, while G. attenuatus has these sclerites V-shaped ( Fig. 5 View FIGURE 5 B). Males of G. tytonis sp. n. are larger on average than G. attenuatus : 360x220 versus 330x200 ( Fig. 7 View FIGURE 7 ). However the shape of epimerites and the body size may sometimes fail as distinguishing characters.
DNA barcode. We amplified and sequenced a 644 bp fragment of the cytochrome oxidase subunit I (COI) gene (DNA barcode region chosen by the Consortium for the Barcode of Life, http://barcoding.si.edu) for one male holotype and 3 male paratypes (Acc. EU271955 View Materials ), remaining paratypes (Acc. EU271956 View Materials ) as well as for specimens of G. attenuatus (Acc. EU271957 View Materials , EU271958 View Materials ). Intraspecific K2P divergence in G. attenuatus was 0.47%. No intraspecific variability in G. tytonis sp. n. was detected. Comparison of the sequences from Glaucalges tytonis sp. n. and G. attenuatus showed that proportion of different nucleotides ranged from 14.65% to 14.85% (mean 14.75%). Because most of the nucleotide substitutions were synonymous, they resulted only in one amino acid change (substitution of valine with isoleucine). This observed genetic distance in the nucleotide sequence of the DNA barcode is substantial, with the differentiation between species comparable to the majority of currently recognized species ( Hebert et al. 2003).
Etymology. The specific epithet is derived from the generic name of the host Tyto alba .
Remarks. Although DNA extraction from a single individual of feather mite is technically possible we suggest in case of small-sized mites such as these Glaucalges species (less than 0.5 mm) to publish DNA barcode sequence isolated from a sample of several individuals (holotype and paratypes = “ type sample”) along with information about possible genetic variability.
Results of the present study seem to confirm our previous observations that multihost feather mite species are often in fact a group of cryptic species. Each of these species is associated with limited host range of closely related bird species or even with a single host species ( Badek & Dabert 2006, Dabert et al. 2005). If this evolutionary scenario holds true, it is likely that the Glaucalges species recorded on other strigid owls may also represent different species of mites. In our opinion, only thorough morphological analysis accompanied by DNA barcoding will significantly accelerate studies that could resolve this and many similar taxonomic problems.
Mite species Hosts Locality References
(Buchholz) Strigidae
Aegolius funereus View in CoL (L.) Germany Eichler (1938) Asio flammeus (Pontoppidan) View in CoL Germany Buchholz (1869) Asio otus View in CoL (L.) Morocco Gaud (1980) Germany Buchholz (1869) Athene noctua (Scopoli) View in CoL Rhodesia, Zaire, Zululand Gaud (1980) Morocco Gaud (1958) Bubo africanus (Temminck) View in CoL Rwanda, Zaire, Zululand Gaud (1980) Bubo bubo View in CoL (L.) Zaire Gaud (1980) Finland Lönnfors (1937) Switzerland Mumcuoglu and Müller (1974) Bubo lacteus (Temminck) View in CoL Rwanda, Zaire Gaud (1980) Bubo leucostictus Hartlaub View in CoL Zaire Gaud (1980) Bubo poensis Fraser View in CoL Morocco Gaud (1958) Zaire Gaud (1980) Cameroon Gaud and Mouchet (1959), Gaud and Till (1961), Gaud (1980) Bubo shelleyi View in CoL (Sharpe et Zaire Gaud (1980) Ussher)
Bubo virginianus (Gmelin) View in CoL USA Atyeo and Philips (1984) Cameroon Gaud (1980) Scotopelia peli (Bonaparte) View in CoL Cameroon Gaud and Till (1961), Gaud (1980) Strix aluco View in CoL (L.) no data Philips (2007) Strix woodfordii (Smith A.) View in CoL Cameroon Gaud (1980)
G. ty t on is sp. n. 2 Tytonidae View in CoL
Tyto alba (Scopoli) Poland, Germany present study
Morocco Gaud and Petitot (1948), Gaud (1958)
Senegal Gaud and Mouchet (1959), Gaud and Till (1961), Gaud (1980) Cameroon, Transvaal Gaud (1980) United Kingdom Rothschild and Clay (1952) Cuba Č erný (1967) Japan McClure and Ratanaworabhan (1973) 1 India Philips (2007)
(Gaud et Mouchet) Musophagidae
Corythaeola cristata (Vieillot)) View in CoL S Cameroon Gaud and Mouchet (1959), Gaud and Atyeo (1981)
1 Designed as Protalges sp. only.
2 Provisional designations without checking the material described under name Glaucalges attenuatus (except present study).


| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |