
Diploria, MILNE EDWARDS & HAIME, 1848: 493
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00855.x |
|
persistent identifier |
https://treatment.plazi.org/id/B26DA91C-6D03-1727-E3F8-FF71D51AF924 |
|
treatment provided by |
Marcus |
|
scientific name |
Diploria |
| status |
|
GENUS DIPLORIA MILNE EDWARDS & HAIME, 1848: 493 View in CoL ( FIGS 10G, H View Figure 10 , 16A, B, 20G–I View Figure 20 , 24G–I View Figure 24 )
Type species: Meandrina cerebriformis de Lamarck, 1816: 246 , holotype = MNHN-Scle102 [= Madrepora labyrinthiformis Linnaeus, 1758: 794–795 ]; by original designation.
Type species locality: ‘Habite les mers d’Amérique’ ( de Lamarck, 1816: 246) [Recent].
Original description: ‘Les séries de polypiérites soudées entres elles par les côtes et l’exothèque qui sont très-développées. Collines doubles et très-larges. Cloisons serrées, très-débordantes et dont les dents supérieures sont les plus fortes.’ ( Milne Edwards & Haime, 1848: 493)
Subsequent morphological descriptions ( Matthai, 1928 and later): Matthai (1928: 50–54); Wells (1936: 118–119, 1956); Vaughan & Wells (1943: 163–164, 166, 171: F402-403); Zlatarski & Estalella (1982: 66–83); Walton Smith (1971: 80–82); Veron (2000: vol. 3: 206–209).
Diagnosis: Macromorphology: meandroid (uniserial), with medium valleys (4–15 mm); well-developed costate coenosteum; continuous, trabecular columella; well-developed epitheca; moderately developed tabular endotheca; small septal lobes ( Figs 10G, H View Figure 10 , 16A, B).
Micromorphology: regular tricorne teeth with pointed tips and elliptical bases orientated transverse to the septal plane; smooth interarea of teeth; spiked granules organized in lines; teeth in different septal cycles and across individual septa similar in size and shape ( Fig. 20G–I View Figure 20 ).
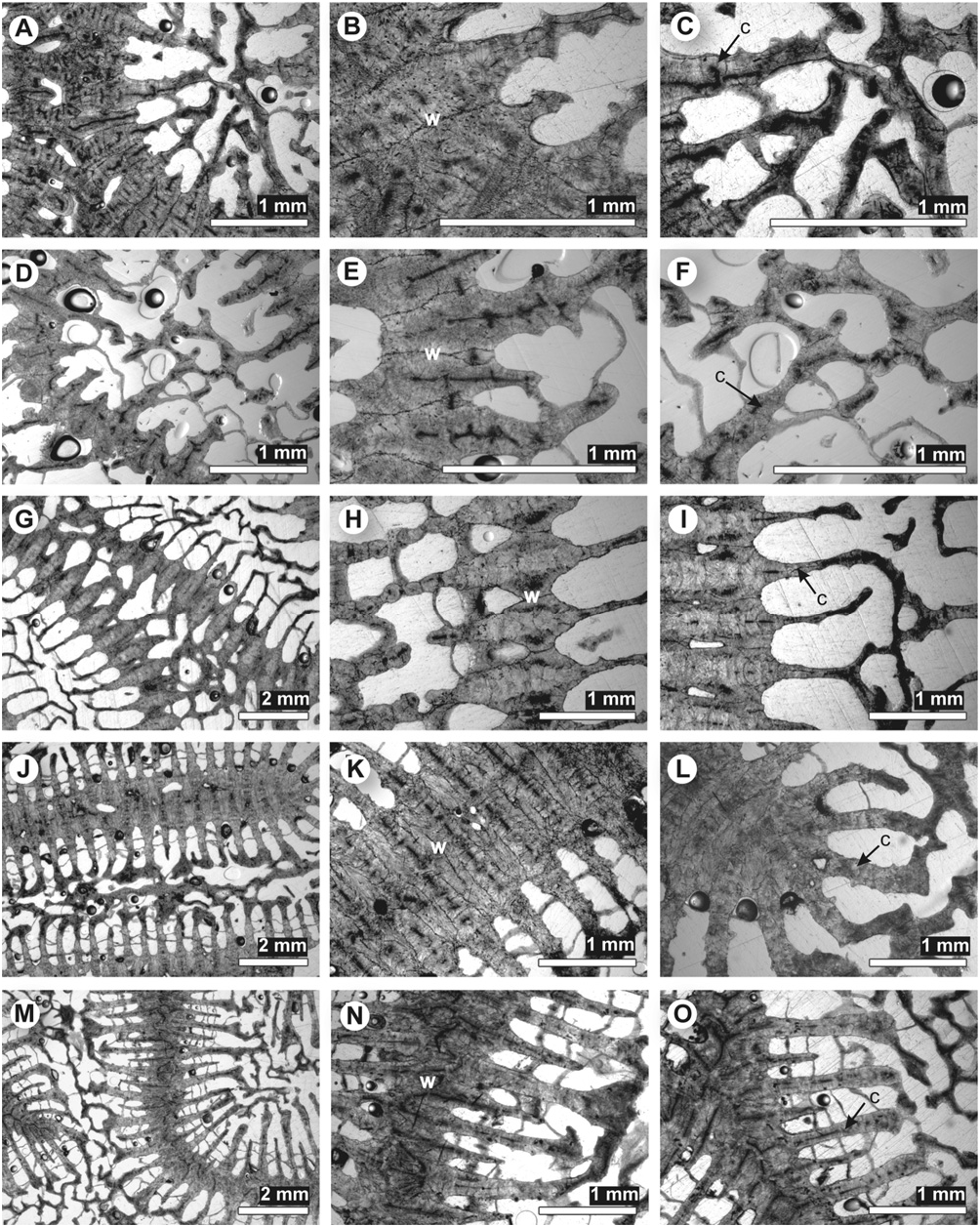
Microstructure: septothecal wall; moderately welldeveloped clusters of calcification centres within costosepta, which are connected by faint or absent medial lines; no carinae; fibrous thickening deposits containing layers ( Fig. 24G–I View Figure 24 ).
Species included: [monotypic genus]: Diploria labyrinthiformis ( Linnaeus, 1758) [holotype is from an unknown locality, and is lost ( Matthai, 1928); neotype (herein designated) = SUI1228814 View Materials ( FA1078 ) collected by H. Fukami in Boca del Toro, Panama, Fig. 16A, B] .
Remarks: Milne Edwards & Haime (1849) originally designated Meandrina cerebriformis as the type species of Diploria based on a specimen in the Lamarck collection (MNHN-Scle102). As indicated in Matthai (1928: 63), this species has subsequently been synonymized with Diploria labyrinthiformis (Linnaeus) , the type specimen of which is lost. A neotype ( SUI 1228814) has been therefore designated for Diploria labyrinthiformis .
In the classification system of Vaughan & Wells (1943) and Wells (1956), the genus Diploria is distinguished by having a meandroid colony form; long sinuous series; a continuous parietal (= trabecular) columella, and a septothecal corallite wall. Septal margins are strongly dentate; trabeculae are usually simple, in one or two fan systems ( Vaughan & Wells, 1943: 153–154, 163, 166). It is distinguished from its meandroid Indo-Pacific counterpart, Platygyra Ehrenberg, 1834 , on the basis of its well-developed columella, septal lobes, septothecal walls (trabeculothecal in Platygyra ), its regular tricorne septal teeth with elliptical-perpendicular bases (irregular multiaxial in Platygyra ), and its well-developed aligned granules.
Cairns, Hoeksema & Land (1999), Zlatarski & Estalella (1982), Walton Smith (1971), and Veron (2000) all recognized three species of Diploria ( Diploria labyrinthiformis , Diploria strigosa , Diploria clivosa ), which differ in valley width, numbers of septal cycles, and development of coenosteum. Diploria labyrinthiformis is distinguished by a welldeveloped coenosteum with a distinctive grooved ambulacrum, valley widths of ~ 5 mm, and 14–17 septa per cm ( Fig. 16A, B). Diploria strigosa has limited coenosteum, valley widths of ~ 6 mm, and 15–20 septa per cm ( Fig. 16C, D). Diploria clivosa has no coenosteum, valley widths of ~ 3.75 mm, and 30–40 septa per cm ( Fig. 16E, F). However, the molecular analyses (nuclear b- tubulin exon and intron; mitochondrial cox1-trnM intergenic region) of Nunes et al. (2008) show that Diploria labyrinthiformis is more closely related to Manicina areolata than to the other two species. Mitochondrial data show Diploria strigosa and Diploria clivosa as grouping together in the same genus-level clade, but nuclear data do not. We therefore separate the three traditional species of Diploria into two genera: one ( Diploria ) containing labyrinthiformis and the other ( Pseudodiploria ) containing strigosa and clivosa . The two genera can be distinguished on the basis of the development of coenosteum, wall structure, and the distinctiveness of medial lines.
GENUS MANICINA EHRENBERG, 1834: 325 View in CoL
( FIGS 11A, B View Figure 11 , 16G–J, 21D–F View Figure 21 , 25D–F View Figure 25 )
Synonyms: Podasteria Ehrenberg, 1834: 326 [type species = Manicina mayori Wells, 1936 (= Manicina gyrosa Ehrenberg, 1834 )]; see Wells, 1936: 125–126.
Type species: Madrepora areolata Linnaeus, 1758: 795 ; by subsequent designation, Milne Edwards & Haime, 1848: 493. Holotype is lost ( Matthai, 1928). We herein designate specimen ZMB-Cni673 from St. Thomas, US Virgin Islands, in the Ehrenberg collection (Berlin Museum) as the neotype. This specimen was identified by Ehrenberg (1834: 324) as ‘ Manicina areolata Enhrenberg’.
Original type species locality: ‘O. Asiatico’ ( Linnaeus, 1758) [Recent].
Original descriptions:
1. ‘Manshetten-Coralle’ ( Ehrenberg, 1834: 325);
2. ‘Se distingue du précédent par son épithèque incomplète et pars ses cloisons (= septa) trèsfortement granulées latéralement, qui offrent près de la columelle un lobe paliforme.’ ( Milne Edwards & Haime, 1848: 493).
Subsequent morphological descriptions ( Matthai, 1928 and later): Matthai (1928: 78–95); Wells (1936: 118–119, 1956); Vaughan & Wells (1943: 163–164, 166, 171: F402-403); Walton Smith (1971: 83–84); Zlatarski & Estalella (1982: 85–92); Veron (2000: vol. 3: 99).
Diagnosis: Macromorphology: meandroid (uniserial), with large valleys (12–15 mm); continuous, spongy trabecular columella; reduced epitheca; tabular endotheca; well-developed septal lobes; sometimes free-living ( Figs 11A, B View Figure 11 , 16G–H).
Micromorphology: tricorne teeth orientated transverse to the septal plane; smooth interarea of teeth; spiked granules organized in lines; columellar teeth similar in size and shape to septal teeth ( Fig. 21D–F View Figure 21 ).
Microstructure: septothecal wall with trabeculothecal elements; well-defined clusters of calcification centres within costosepta, which are connected by medial lines; carinae; fibrous thickening deposits containing layers ( Fig. 25D–F View Figure 25 ).
Species included: Manicina areolata ( Linnaeus, 1758: 795) [holotype is from an unknown locality, and is lost ( Matthai, 1928); neotype (herein designated) = ZMB- Cni 673, Fig. 11A, B View Figure 11 , St. Thomas, US Virgin Islands] .
Manicina mayori Wells, 1936 View in CoL (= Manicina gyrosa Ehrenberg, 1834: 326 View in CoL ); holotype = ZMB-Cni2859, unknown locality), Fig. 16I, J (also figured by Matthai, 1928: pl. 63, fig. 6).
Not Madrepora gyrosa Ellis & Solander, 1786 [= Colpophyllia natans ( Houttuyn, 1772) View in CoL ].
Remarks: In the classification system of Vaughan & Wells (1943) and Wells (1956), the genus Manicina is distinguished by ‘reduced epitheca, broader and more open corallum, longer series, usually continuous, with ambulacra. Inner septal lobes small and narrow’ ( Vaughan & Wells, 1943: 171). The genus Diploria is ‘meandroid..., forming massive or subencrusting colonies with long series and thick collines with narrow or broad ambulacra. Septa of some species with small internal lobes. Columella continuous, parietal’ ( Vaughan & Wells, 1943: 166). In the key to genera ( Vaughan & Wells, 1943: 162), the two genera are distinguished by the presence of internal lobes ( Manicina : present; Diploria : absent) and by corallite wall structure ( Manicina : ‘parathecal’; Diploria : septothecal). Contrary to these distinctions, our examination showed that Diploria labyrinthiformis and the two species of Pseudodiploria ( Pseudodiploria strigosa , Pseudodiploria clivosa ) sometimes have small septal lobes (like Manicina ) and the wall of the two Pseudodiploria species is partially trabeculothecal (like Manicina ). Both characters are therefore highly variable, as are other characters that have been cited as diagnostic, including valley width and development of epitheca. Similar macromorphological characters include (1) uniserial, meandroid form; (2) continuous, trabecular columella; and (3) tabular endotheca. Our study of microstructural and micromorphological characters shows that Manicina , Diploria , and Pseudodiploria have similar septal teeth and interareas between teeth. However, in contrast to Diploria and to a lesser extent Pseudodiploria , Manicina forms distinct clusters of calcification centres that are connected by medial lines, carinae are present, septal teeth are especially well developed, and the columella is spongy.
Cairns, Hoeksema & Land (1999), Zlatarski & Estalella (1982), and Veron (2000) only recognized one species of Manicina , Manicina areolata ; whereas Walton Smith (1971), following Matthai (1928) and Wells (1936), recognized two species, Manicina areolata and Manicina mayori , which are distinguished on the basis of colony form. Manicina areolata forms small, free-living colonies composed of one continuous valley ( Fig. 16G, H), whereas Manicina mayori has larger, attached colonies with many discontinuous valleys ( Fig. 16I, J).
GENUS MUSSISMILIA ORTMANN, 1890: 292 View in CoL
( FIGS 11C–F View Figure 11 , 17A–H View Figure 17 , 21G–O View Figure 21 , 25G–O View Figure 25 )
Synonyms: Protomussa Matthai, 1928: 268 [type species = Acanthastraea braziliensis Verrill, 1868: 355 ].
Type species: Mussa harttii Verrill, 1868: 357 , syntypes = YPM1468 View Materials A-C, 1469; by original designation. We herein designate YPM1468 View Materials A collected by C. F. Hartt (1867) on Abrolhos Reef in Brazil, as the lectotype of Mussismilia hartti (Verrill) .
Type species locality: Abrolhos Reef , Abrolhos Archipelago, Brazil [Recent] .
Original description: ‘Dieses Verhalten macht es nothwendig, die Mussa harti von der Gattung Mussa zu trennen and dieselbe den folgenden Formen anzuschiessen. Wegen der eigenthümlichen, stark bedornten Rippen wird sie wohl eine eigene Gattung bilden müssen, für die ich die Namen Mussismilia vorschlagen möchte’ ( Ortmann, 1890: 292).
Subsequent morphological descriptions ( Matthai, 1928 and later): = Protomussa in Matthai (1928: 268– 272); Vaughan & Wells (1943: 192–193); Wells (1956: F417); Laborel (1969: 195–198); Veron (2000: vol. 3: 118); Walton Smith (1971: 80).
Diagnosis: Macromorphology: short series (one to three centres per series); discontinuous columella with trabecular linkage; abundant vesicular endotheca; no paliform lobes ( Figs 11C–F View Figure 11 , 17A–H View Figure 17 ).
Micromorphology: regular lacerate (twisted threads; multicentred, multidirectional) teeth; evenly scattered, spine-shaped septal granulation; porous septa; teeth in major and minor septal cycles are equal in size and shape; spongy columella, with columellar teeth different in size and shape from septal teeth ( Fig. 21G–O View Figure 21 ).
Microstructure: parathecal (dissepiments only) corallite wall, sometimes containing trabeculothecal elements; centres of calcification within the wall, septa, and columella form a medial line broken by distinct clusters of centres; fibrous thickening deposits containing layers ( Fig. 25G–O View Figure 25 ).
Included species: Mussismilia hartti ( Verrill, 1868: 357) [lectotype = YPM1468 View Materials A, Fig. 11C–F View Figure 11 , Abrolhos , Brazil] .
Mussismilia braziliensis ( Verrill, 1868: 355) View in CoL [holotype = YPM1467 View Materials , Fig. 17C, D View Figure 17 , Abrolhos , Brazil] .
Mussismilia hispida ( Verrill, 1901: 127) View in CoL [holotype = YPM4287 View Materials , Fig. 17E, F View Figure 17 , Abrolhos , Brazil] .
Mussismilia leptophylla ( Verrill, 1868: 353) View in CoL [holotype = YPM1517 View Materials A, Fig. 17G, H View Figure 17 , Abrolhos, Brazil] (= Heliastraea aperta Verrill, 1868: 356 ) [holotype = YPM1518 View Materials , Abrolhos , Brazil] .
Remarks: In the classification system of Vaughan & Wells (1943) and Wells (1956), the genus Mussismilia was placed within the family Mussidae because of its ‘ragged’ septal dentations and because its septa are formed by several (> two) trabecular fan systems. Our examination of micromorphology shows that the shape of its septal teeth is indeed unique. The teeth are medium to high and formed by multiple twisted threads, which are uniform in arrangement but vary in size and orientation. The septa are usually thin and often porous near the columella. In addition, the genus differs from Manicina and Favia in its abundant vesicular endotheca, its parathecal (dissepiments only) wall, its costoseptal microstructure (welldeveloped clusters of calcification centres), and its interarea of septal teeth (horizontally banded). The genus is currently restricted to Brazil, but was common across the Caribbean region during the Miocene and Pliocene ( Budd, Stemann & Johnson 1994; Budd, Petersen & McNeill, 1998; Budd et al., 1999).
In the classification system of Vaughan & Wells (1943) and Wells (1956), the species Mussismilia leptophylla was placed in the genus Favia , presumably because of the lack of multiple trabecular fan systems. However, our observations indicate that Mussismilia leptophylla also has more than one fan system ( Fig. 21M–O View Figure 21 ). In addition, its parathecal wall structure, septal microstructure, and the shape of its septal teeth and granules are similar to those of Mussismilia . Moreover, based on nuclear and mitochondrial markers, Nunes et al. (2008) found that Mussismilia leptophylla groups more closely with the three Mussismilia species than it does with Favia fragum or Favia gravida . We therefore transfer Mussismilia leptophylla for the first time to the genus Mussismilia .
Cairns, Hoeksema & Land (1999) and Veron (2000) recognized three species of Mussismilia , which differ in colony form, calice size, and septal thickness (as does Mussismilia leptophylla , per our observations). Mussismilia hartti has a phaceloid colony form, calice diameters of 12–30 mm, 12–14 septa per cm, and thin septa ( Fig. 17A, B View Figure 17 ); whereas Mussismilia hispida , Mussismilia braziliensis , and Mussismilia leptophylla are all subplocoid. Mussismilia hispida has calice diameters of 14–15 mm, more than four septal cycles, thin septa, and a well-developed columella ( Fig. 17E, F View Figure 17 ); Mussismilia braziliensis has calice diameters of 8–10 mm, fewer than four septal cycles, thick septa, and a weak columella ( Fig. 17C, D View Figure 17 ); and Mussismilia eptophylla has calice diameters of ~ 6 mm, fewer than four septal cycles, thin septa, and a well-developed columella ( Fig. 17G, H View Figure 17 ).
GENUS PSEUDODIPLORIA FUKAMI, BUDD & KNOWLTON GEN. NOV. ( FIGS 11G, H View Figure 11 , 16C–F, 20J–O View Figure 20 , 24J–O View Figure 24 )
Synonyms: Maeandrina Link, 1807 , sensu Matthai, 1928: 50–54;
Type species: Meandrina strigosa Dana, 1846: 257–258 ; pl. 14, fig. 4a–b; holotype = USNM00005 (= Meandrina cerebrum Ellis & Solander, 1786: 163 ; Matthai, 1928: 55–63).
Type species locality: ‘ West Indies’ ( Dana, 1846) [Recent] .
Diagnosis: Macromorphology: meandroid (uniserial), with medium valleys (4–15 mm); little or no coenosteum; continuous, trabecular columella; welldeveloped epitheca; moderately developed tabular endotheca; small septal lobes ( Fig. 11G, H View Figure 11 , 16C–F).
Micromorphology: regular tricorne teeth with pointed tips and elliptical bases orientated transverse to the septal plane; smooth interarea of teeth; spiked granules organized in lines; teeth in different septal cycles and across individual septa similar in size and shape ( Fig. 20J–O View Figure 20 ).
Microstructure: septothecal wall with trabeculothecal elements; weak clusters of calcification centres within costosepta, which are connected by medial lines; no carinae; fibrous thickening deposits containing layers ( Fig. 24J–O View Figure 24 ).
Species included: Pseudodiploria strigosa ( Dana, 1846: 257–258; pl. 14, fig. 4a–b) [holotype = USNM00005 View Materials , Fig. 11G, H View Figure 11 , West Indies ] [= Maeandrina cerebrum (Ellis & Solander) of Matthai, 1928] .
Pseudodiploria clivosa ( Ellis & Solander, 1786: 163) View in CoL [holotype is from ‘Oceano Indiae occidentalis’, and is lost ( Matthai, 1928); neotype (herein designated) = SUI122811 View Materials , Fig. 16E, F, Bocas del Toro, Panama] .
Remarks: Matthai (1928) indicated that Maeandrina cerebrum (Ellis & Solander) is the type species of Maeandrina Link, 1807 . However, given that Meandrina de Lamarck, 1801 predates Maeandrina Link, 1807 and is a valid genus name ( Table 1, Family Meandrinidae ), we herein name a new genus, Pseudodiploria .
As explained above under Diploria , Pseudodiploria can be distinguished from Diploria by its limited or absent coenosteum, the presence of trabeculothecal elements in the otherwise septothecal corallite wall, and its often well-developed medial lines. The two species of Pseudodiploria ( Fig. 16C–F) differ in the presence of coenosteum ( Pseudodiploria strigosa has limited coenosteum but Pseudodiploria clivosa has no coenosteum), valley width ( Pseudodiploria strigosa has valley widths of ~ 6 mm but Pseudodiploria clivosa has valley widths of ~ 3.75 mm), and number of septa per cm ( Pseudodiploria strigosa has 15–20 but P. clivosa has 30–40 septa per cm).






| SUI |
The University of Iowa (formerly State University of Iowa) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Diploria
| Budd, Ann F., Fukami, Hironobu, Smith, Nathan D. & Knowlton, Nancy 2012 |
Mussismilia hispida ( Verrill, 1901: 127 )
| Verrill AE 1901: 127 |
MUSSISMILIA ORTMANN, 1890: 292
| Ortmann AE 1890: 292 |
Mussismilia braziliensis ( Verrill, 1868: 355 )
| Verrill AE 1868: 355 |
Mussismilia leptophylla (
| Verrill AE 1868: 353 |
| Verrill AE 1868: 356 |
DIPLORIA MILNE EDWARDS & HAIME, 1848: 493
| Milne Edwards H & Haime J 1848: 493 |
MANICINA EHRENBERG, 1834: 325
| Ehrenberg CG 1834: 325 |
Manicina mayori
| Ehrenberg CG 1834: 326 |
Pseudodiploria clivosa ( Ellis & Solander, 1786: 163 )
| Ellis J & Solander DC 1786: 163 |