
Lochmanolenellus trapezoidalis, Webster, Mark & Bohach, Lisa L., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3824.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:023D78D0-4182-48D2-BAEB-CDA6473CF585 |
|
DOI |
https://doi.org/10.5281/zenodo.6129738 |
|
persistent identifier |
https://treatment.plazi.org/id/B10C8793-FFC8-FFB5-61B5-FEB8FAE4806C |
|
treatment provided by |
Plazi |
|
scientific name |
Lochmanolenellus trapezoidalis |
| status |
sp. nov. |
Lochmanolenellus trapezoidalis n. sp.
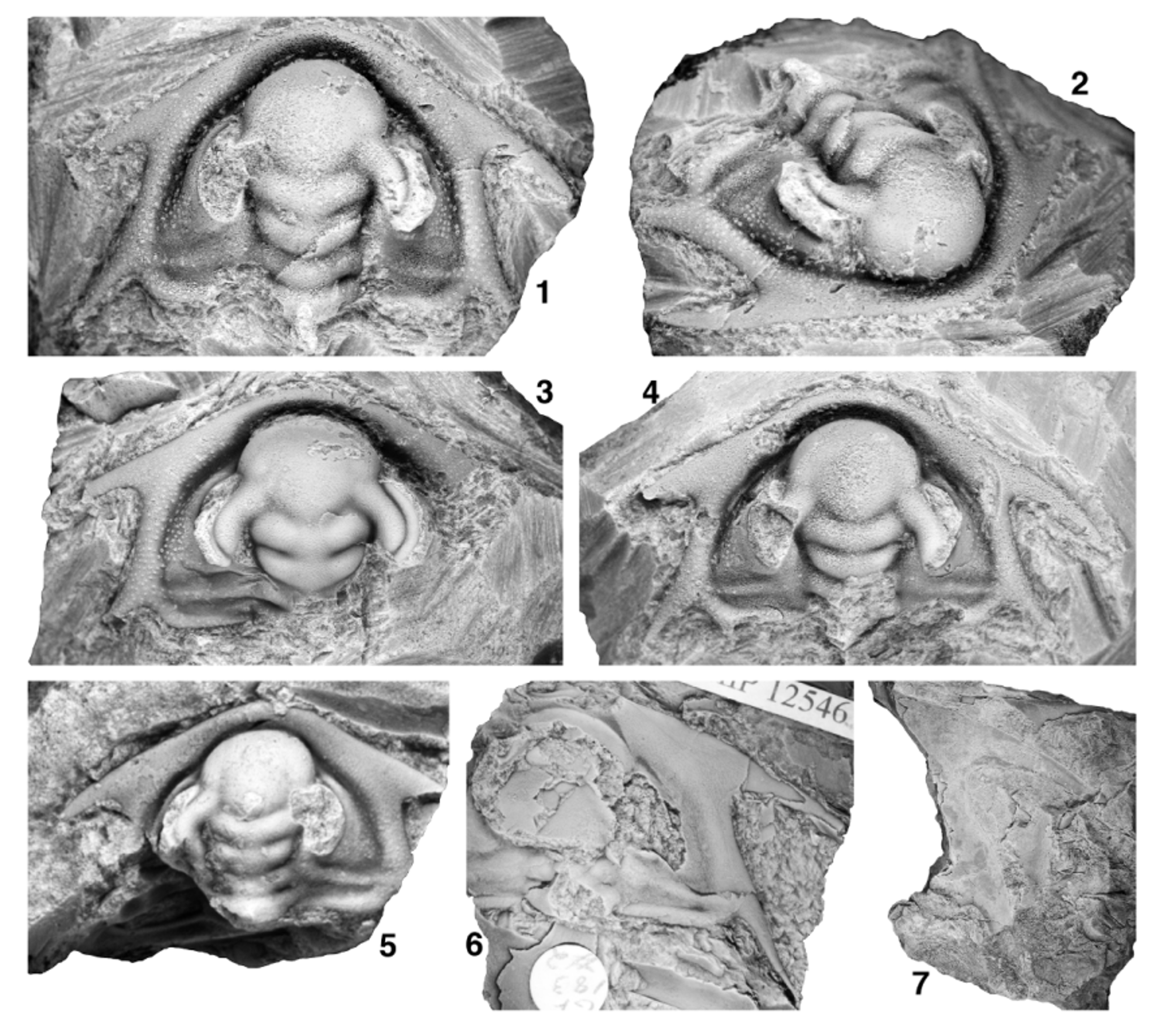
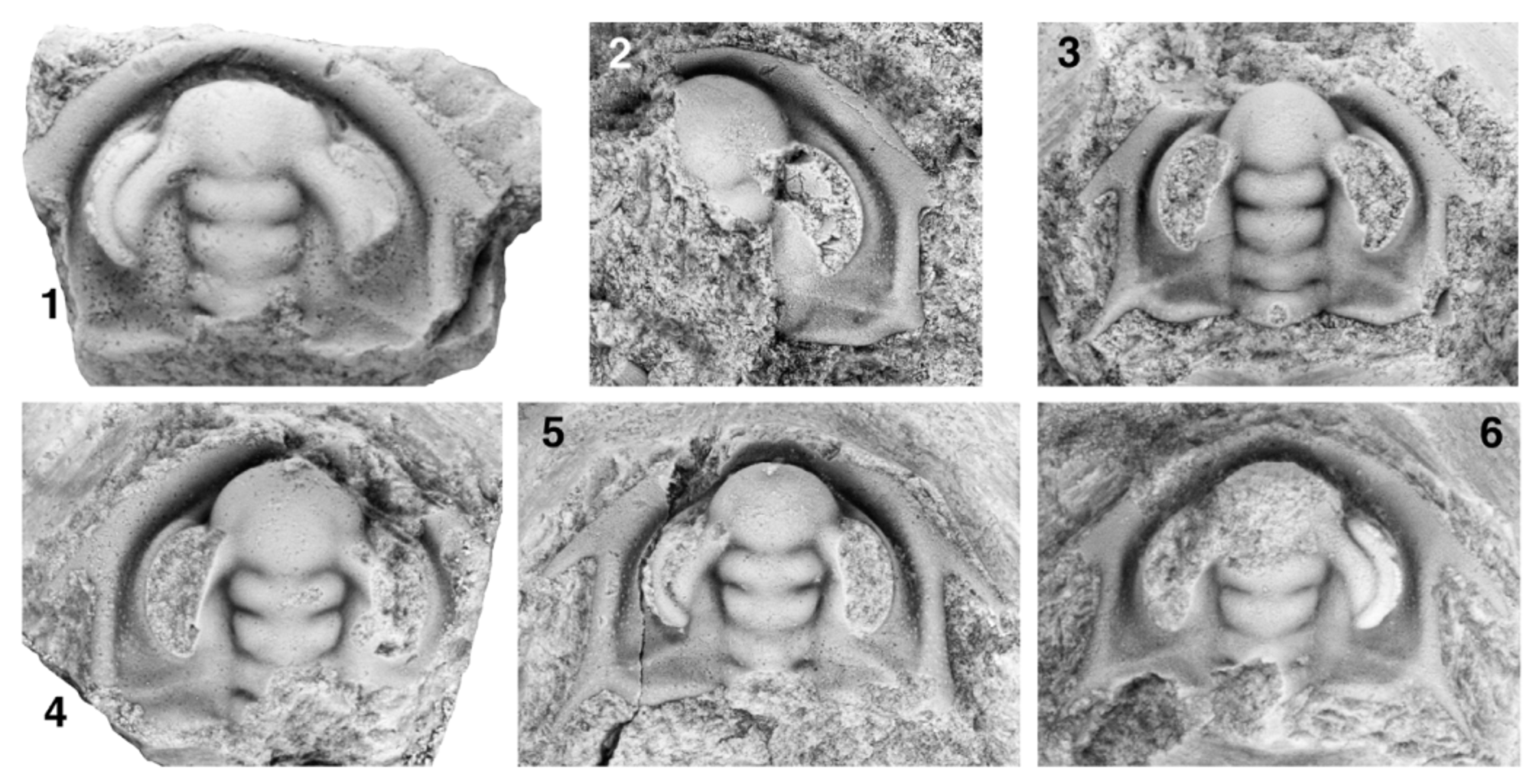
Figs 20 View FIGURE 20. 1 – 9 , 21 View FIGURE 21 , 22 View FIGURE 22
1976 Laudonia sp.; Nelson (part), pl. 5, top left “ Laudonia ” specimen only [not top right or lower left “ Laudonia ” specimens, = Lochmanolenellus cf. subquadratus 1].
Diagnosis. Cephalon trapezoidal in outline; genal spine bases located transversely opposite posterior portion of LA (sag.); distal portion of posterior cephalic margin oriented anteriorly and slightly inwards when traced toward base of genal spine. Posterior tip of ocular lobe transversely opposite distal tip of S1 or lateral margin of L2.
Description (mature morphology, phase 5 of cephalic development; sagittal cephalic length> 4.5 mm)
Cephalon trapezoidal in outline, widest (tr.) at base of intergenal spines. Cephalic length of largest known specimen estimated to have been approximately 35 mm (sag.; Fig. 20.8, 20.9). Proximal portion of posterior cephalic margin oriented weakly posteriorly by up to 8° relative to a transverse line when traced abaxially, flexing anteriorly at a rounded adgenal angle located slightly less than two-fifths distance along cephalic margin from axial furrow to base of genal spine, distal portion of posterior cephalic margin oriented anteriorly and inwards at 7° to 19° (increasing through phase 5) relative to an exsagittal line when traced toward base of genal spine. Genal spine stout, broad-based, tapers to point; angle between distal portion of posterior cephalic margin and inner margin of proximal portion of spine approximately 49° to 67° (generally increasing through phase 5), spines very slightly curving into more posterior orientation along their length; spine length slightly less than one-half cephalic length (sag.); spine base at contact with distal portion of posterior cephalic margin transversely opposite posterior portion of LA (sag.; anterior to anterior-most point of S3). Distance (tr.) between genal spine bases approximately 110% to 135% of cephalic length (sag.). Intergenal spine slender, broad-based, angle between distal portion of posterior cephalic margin and outer margin of proximal portion of spine approximately 157° to 179°; tips rarely preserved but spine length estimated to be slightly shorter than genal spine; spine base slightly distal to adgenal angle. Cephalic border defined by broad, trough-like border furrow interrupted only by intergenal ridge as it merges with border at base of intergenal spine; border furrow somewhat shallower and narrower along proximal portion of posterior cephalic border. Anterior cephalic border narrowest anterior to LA, broadens distally, width opposite junction of ocular lobes with LA approximately 13% to 25% length (sag.) of glabella (increasing though phase 5); broadest portion of border located at confluence with genal spines; posterior border tapers to point adaxially, barely touches axial furrow at posterior-most portion of LO. Border well rounded dorsally anterior to LA, somewhat more broadly dorsally arched at and between bases of genal and intergenal spines. Preglabellar field absent; LA slopes anteriorly almost vertically into or slightly overhangs trough-like cephalic border furrow that is slightly wider than anterior border at sagittal axis. Plectrum not developed. Glabella club-shaped in outline, most constricted (tr.) at L1, 86% to 91% of cephalic length (sag.), moderately dorsally convex (tr.), strongly dorsally arched (sag.), summit formed by posterior portion of LA and L3. Maximum width of LA 134% to 155% basal glabellar width (tr.). Posterior margin of glabella convex posteriorly. SO transglabellar but deepest abaxially, abaxial end slightly anterior to adaxial end. LO subquadrate (transverse width across SO averages approximately 101% [range 96% to 106%] of basal glabellar width), length (exsag.) approximately 9% to 12% of glabellar length (sag.), very strongly dorsally convex (tr.). S1 transglabellar but deepest abaxially, approximately parallel to SO, slightly deeper than SO over axis. L1 weakly trapezoidal (narrowing slightly anteriorly) to subquadrate, transverse width across S1 approximately 93% (range 87% to 100%) of basal glabellar width; length (exsag.) approximately 10% to 15% of glabellar length (sag.). S2 transglabellar but deepest abaxially, connected to axial furrow, strongly arcuate (convex anteriorly) on either side of sagittal axis, adaxial portion slightly to well posterior to distal portion. L2 broadly Vshaped, lateral margins strongly diverging anteriorly, transverse width across S2 approximately 110% (range 100% to 118%) of basal glabellar width, distance (exsag.) between contact of S1 with axial furrow and contact of S2 with axial furrow approximately 13% of glabellar length (sag.). S3 transglabellar, oriented strongly anterolaterally away from axis until contact with ocular lobes where it contacts anteriorly converging axial furrow, the two together thus forming a strongly caret-shaped furrow on either side of sagittal axis, deepest at apex of each caret. L3 broadly Mshaped, lateral margins diverging anteriorly until contact with inner margin of ocular lobes, then converging anteriorly until contact with S3; transverse width of glabella at point of contact between axial furrow and inner margin of ocular lobes approximately 121% (range 112% to 128%) of basal glabellar width, distance (exsag.) between contact of S2 with axial furrow and contact of axial furrow with inner margin of ocular lobes approximately 6% (range 4% to 8%) of glabellar length (sag.). LA impinges into and anteriorly deflects anterior border furrow; transversely oblate in outline, anterior margin broadly rounded; maximum width (tr.) typically at contact with outer margin of ocular lobes (rarely slightly anterior to that; Fig. 20.7), length (sag.) approximately 77% (range 68% to 84%) of maximum width and 41% to 49% of glabellar length (sag.), maximum width (tr.) approximately 146% (range 135% to 155%) of basal glabellar width; moderately dorsally convex (tr.), strongly dorsally convex (sag.); prominently inflated relative to extraocular area, lateral margins clearly defined from anterior portion of extraocular area by axial furrow. Extremely shallow, weakly arcuate (convex posteriorly) preocular furrow runs inwards and anteriorly from contact of LA with outer margin of ocular lobes; not incised over axis. Very shallow transocular furrow runs anterolaterally across ocular lobes from point of contact between S3 and axial furrow to preocular furrow, isolating ocular lobes from LA. Axis of LO often chipped, but preserves base of prominent axial node or short axial spine. Subdued ovoid lateral swellings on L2. Anterolateral portion of LO merges into interocular area, interrupting axial furrow. Axial furrow broad, deepest at lateral margins of L2 and L3. Ocular lobes divergent from exsagittal axis by approximately 31° (range 20° to 40°), crescentic; posterior tip transversely opposite distal tip of S1 (Fig. 21.4) to anterior portion of lateral margin of L2 (more anteriorly located on larger specimens; Fig. 20.1–20.7). Deep ocular furrow runs along length of ocular lobe slightly distal to midline so that inner band of ocular lobe is wider (tr.) than outer band; anteriorly, ocular furrow curves slightly outwards to merge into a “triple junction” of furrows at point where preocular furrow meets the axial furrow at the margin of LA, thus isolating outer band of ocular lobe from LA; posteriorly, ocular furrow curves slightly inwards to meet inner margin of ocular lobe so that posterior tip of ocular lobe is located on outer band. Inner and outer bands of ocular lobe each convex dorsally (tr.). Circumocular suture defines large visual surface (not preserved) that occupied much of length of outer wall of ocular lobe; portion of outer wall of ocular lobe below circumocular suture forms prominent, steep eye socle approximately one-quarter of estimated vertical height of visual surface. Interocular area slopes down from inner margin of ocular lobe to axial furrow; approximately half width (tr.) of inner band of ocular lobe opposite S2. Extraocular area climbs moderately steeply out of broad border furrow then arches to produce a moderately vaulted transverse profile. In vicinity of ocular lobe, extraocular area abruptly changes slope to form a steep-walled, narrow-topped (tr.) extraocular platform upon which ocular lobe sits; extraocular platform subcrescentic in outline when viewed from above; anteriorly, extraocular platform narrows to a thin, subdued ridge that extends beyond anterior limit of ocular lobe, descending in vertical height and running parallel to lateral margin of LA then fading into anterior border furrow; posteriorly, extraocular platform terminates just posterior to posterior tip of ocular lobe. Prominent intergenal ridge extends from interocular area opposite lateral margin of L1 to posterior cephalic border at base of intergenal spine; ridge slightly narrows distally and merges smoothly into border, interrupting border furrow. Network of raised ridges forming walls of polygonal, mesh-like prosopon on dorsal surface of extraocular area, cephalic border furrow, cephalic border, intergenal ridge, bases of intergenal and genal spines (Figs 20.1–20.9, 21.1–21.5), and sometimes interocular area, ocular lobes, and glabella (Figs 20.1–20.6, 21.3); polygons become more elongate (long axes parallel to outer wall of ocular lobe) on extraocular platform, more elongate (long axes parallel to axial furrow) on LA, and more elongate (long axes parallel to cephalic margin) on outer portion of distal posterior cephalic border and anterior cephalic border, often grading into bertillon-like pattern of anastomosing ridges on outermost cephalic border and genal and intergenal spine bases that in turn grades into subparallel lirae on sagittal portion of anterior cephalic border and distal portions of genal and intergenal spines (Figs 20.5, 20.9, 21.6). Central raised boss developed in most polygons; central bosses developed into large, pustule-like structures in some polygons on distal portion of extraocular area, in cephalic border furrow, on intergenal ridge, on posterior cephalic border (particularly on inner portions), and on base of intergenal spines (Fig. 20.5, 20.9). Tropidium-like ridge developed in cephalic border furrow on either side of cephalon adjacent to genal spine bases on one specimen (Fig. 21.4); ridge fades anteriorly and posteriorly into border furrow.
Mature hypostome (Fig. 20.10–20.13) moderately convex (tr., sag.), subtrapezoidal (widest anteriorly) in outline. Anterior lobe of middle body subtriangular, occupies approximately 85% of sagittal length of hypostome; anterior margin broadly curved; extends laterally onto broad, triangular anterior wings, tips of which are located slightly anterior to hypostomal midlength. Posterior lobe of middle body very narrow, <10% of hypostome length (sag.), transversely crescentic in outline, widest anteriorly, divided from anterior lobe over sagittal axis by broad, shallow furrow. Maculae relatively deep. Anterior border very narrow, rim-like; defined by shallow furrow that broadens and deepens abaxially. Lateral and posterior border narrow, approximately 6% of hypostome length sagittally, weakly defined by shallow furrow. Six pairs of small spines/nubbins project posterolaterally from margin of lateral border; abaxial-most to fifth pair form a size gradient from nubbins to small spines, sixth (innermost) pair nubbin-like and almost absent on largest known hypostome (Fig. 20.13); posterior border more-or-less straight (tr.) and without spines. Network of raised ridges forming walls of polygonal, mesh-like prosopon on ventral surface of anterior and posterior lobes; central raised boss developed in most polygons. Rostral plate, thorax, and pygidium unknown.
Ontogeny. Specimens examined herein range from approximately 2.5 mm to 35 mm in cephalic length (sag.), offering insight into mid- to late portions of cephalic ontogeny. Specimens of sagittal cephalic length> 4.5 mm are essentially morphologically mature and have been described previously.
On the smallest known cephalon (Fig. 22.1) the glabella is more-or-less parallel-sided between SO and S2 (LO is not preserved); L3 is slightly wider (tr.) than L2, and the axial furrow at the anterolateral margins of L3 runs confluent against the inner margin of the base of the ocular lobe. This glabellar morphology is typical of early phase 4 of cephalic development (described previously). L2 and L3 are each slightly proportionally longer (exsag.; approximately 16% and 10% of sagittal glabellar length, respectively), and LA is proportionally much shorter (sag.; 36% of sagittal glabellar length and 59% of the maximum transverse width of LA) than at morphological maturity. The ocular lobes of this specimen are proportionally relatively long, with the posterior tips being transversely opposite the lateral margins of the anterior-most portion of L1. The cephalon is pentagonal rather than trapezoidal in outline; the bases of the genal spines are transversely opposite the distal tip of S2, and the distal portion of the posterior cephalic margin seems to run almost parallel to an exsagittal line (although this cannot be reliably quantified because the posterolateral portions of the cephalon are not preserved). The extraocular platform is already present at this stage of ontogeny. The interocular area is more shelf-like and does not slope as steeply down (tr.) from the ocular lobes to the axial furrow. The polygonal network of raised ridges forming the cephalic prosopon is not evident on this specimen. However, a slightly larger but damaged and exfoliated specimen (Fig. 22.2, estimated cephalic length [sag.] approximately 2.9 mm) clearly shows pustule-like structures on the distal extraocular area that, by comparison to non-exfoliated specimens, are known to be enlarged central bosses of the polygonal prosopon. This specimen also bears procranidial spines, the presence or absence of which cannot be unambiguously determined on the smaller cephalon to due the worn anterior cephalic margin.
Four slightly larger specimens ranging up to approximately 3.6 mm in sagittal cephalic length (Fig. 22.3–22.6) show a glabellar morphology typical of earliest phase 5 of cephalic development—both L2 and L3 widen anteriorly from S1 to the contact of the lateral margins of L3 with the inner margin of the ocular lobe so that the glabella is most constricted (tr.) at S1. (Entry into phase 5 of cephalic development is therefore estimated to have been achieved at sagittal cephalic length of approximately 2.7 mm.) When arranged in order of cephalic size, these specimens also document a transition from a subpentagonal cephalic outline to a subtrapezoidal cephalic outline—the proximal portion of the posterior cephalic margin (traced from the axial furrow to the adgenal angle) progressively proportionally widens (tr.), and the distal portion of the posterior cephalic margin (traced from the intergenal to the genal spine bases) progressively rotates inward so that the cephalon narrows (tr.) from the intergenal spine bases to the genal spine bases. The genal spine bases progressively migrate anteriorly to a position transversely opposite the lateral margins of the anterior-most portion of L3. The ocular lobes progressively proportionally shorten very slightly, with the posterior tips migrating anteriorly from a position transversely opposite the lateral margin of the anterior-most portion of L1 to a position more-or-less opposite the distal tip of S1. The interocular area progressively tilts to slope down (tr.) from the ocular lobes to the axial furrow. “Pustules”, presumably representing the enlarged central bosses of the polygons of the mesh-like prosopon (see previous), are apparent on the extraocular area, intergenal ridge, cephalic border, and intergenal spine bases at this stage (Fig. 22.4–22.6), and the polygonal network of raised ridges is visible on the extraocular area of the largest specimen during this portion of ontogeny (Fig. 22.6).
Other trends in morphology over the sampled portion of ontogeny (early phase 4 to phase 5) include: (1) A marked progressive increase in the proportional length of LA relative to the maximum transverse width of LA (Fig. 23.1) and to glabellar length (sag.; Fig. 23.2). This proportional elongation of LA is partially accommodated by a slight decrease in the proportional length (exsag.) of L1 and L2 relative to glabellar length throughout ontogeny (Fig. 23.3) and a slight decrease in the proportional length (exsag.) of L3 relative to glabellar length until cephalic length of approximately 4.5 mm (beyond which the proportional length of L3 stabilizes; Fig. 23.4). (The proportional length [exsag.] of LO relative to glabellar length does not change over the sampled portion of ontogeny.) However, such accommodation is incomplete because the elongation of LA causes a slight increase in proportional length of the glabella relative to cephalic length over the sampled portion of ontogeny (sag., Fig. 23.5) as LA impinges into and deflects the anterior border furrow. (2) A proportional lateral expansion (tr.) of LA, L3 and L2 relative to L1 throughout phase 5 of cephalic development. (3) A marked proportional shortening of the ocular lobes relative to glabellar length (Fig. 23.6), such that the posterior tips of the ocular lobes migrate anteriorly to a position opposite the anterior portion of the lateral margins of L2 on the largest cephala. This results in a general increase in the tightness of curvature of the outer margin of the ocular lobes. (4) A marked increase in the proportional width of the anterior cephalic border (Fig. 23.7). (5) Anterior migration of the genal spine bases (Fig. 23.8), such that they are located transversely opposite the posterior portion of LA (sag.) on specimens> 4.5 mm in sagittal cephalic length. (6) Progressive inward rotation of the distal portion of the posterior cephalic margin (traced from the intergenal to the genal spine bases; Fig. 24.1). This also results in the distal portion of the posterior cephalic margin becoming increasingly more parallel with the orientation of the intergenal spines (Fig. 24.2). (7) A marked proportional increase in the length of the proximal portion of the posterior cephalic margin, resulting in an increase in the proportional transverse distance between the adgenal angles and intergenal spine bases (Fig. 24.3) relative to glabellar length. Concomitant with this elongation, the proximal portion of the posterior cephalic margin becomes oriented less strongly posterolaterally when traced distally (Fig. 24.4). Trends (5) and (6) result in the genal spines appearing to ‘flare out’ more strongly from the sides of the cephalon (Fig. 24.5). Trends (5), (6), and (7) result in the cephalon becoming increasingly trapezoidal in outline (i.e., narrowing [tr.] anteriorly from the intergenal spine bases to the genal spine bases; Fig. 24.6).
Etymology. Named for the trapezoidal cephalic outline.
Holotype. GSC 137502 (Fig. 20.1–20.5), from GSC locality 95191, Cassiar Mountains, British Columbia.
Other material. Quantitative morphometric and/or qualitative descriptive data were recorded from first-hand examination of the holotype (see previous) and the following 23 cephala: 19 cephala from GSC locality 95191 (including GSC 137487, GSC 137503 to GSC 137505, GSC 137509 to GSC 137512, and GSC 137514 to GSC 137517); one cephalon from GSC locality 95192 ( GSC 137513); one fragmentary cephalon from ICS-1336 ( FMNH PE 58490; no morphometric data taken); and two cephala from LACMIP locality 17041 (LACMIP 12545 and LACMIP 12546). One of the three specimens illustrated and identified as “ Laudonia ” by Nelson (1976, pl. 5, top left specimen only) shows the trapezoidal cephalic outline, strongly anteriorly advanced genal spine bases, and short ocular lobes typical of Lochmanolenellus trapezoidalis and is herein reassigned to that species. Unfortunately, the whereabouts of that specimen is unknown (M. Stecheson, personal communication to M. Webster, March 2012), and it therefore was not examined during the course of the present study. Three hypostomes from GSC locality 95191 ( GSC 137506 to GSC 137508) are also assigned to this species.
Occurrence. CANADA: Cassiar Mountains, British Columbia: GSC locality 95191, from a thin, wavybedded limestone approximately 18.75 metres above the base of Unit 4 of the Rosella Formation; and GSC locality 95192, from a limestone approximately 25 metres above the base of Unit 4 of the Rosella Formation; both in Section 2 of Fritz (1978, text-fig. 3.1a), on the north bank of the Dease River, approximately 2.7 km ( 1.7 miles) east of McDame. U.S.A.: Montezuma Range, Esmeralda County , Nevada: LACMIP locality 17041, grey to orange mottled packstone approximately 251 metres above the base of the section, corresponding to approximately 183 metres above the base and 18 metres below the top of the middle member of the Poleta Formation ( Bohach, 1997, text-fig. 60, p. 369); and ICS-1336, a bioclastic limestone from an unconstrained stratigraphic position in the lower Dyeran portion of the middle member of the Poleta Formation; both from the IS-4 section of Hollingsworth (1999), Indian Springs Canyon. The specimen illustrated by Nelson (1976, pl. 5, top left specimen) was collected from the middle member of the Poleta Formation in either Esmeralda County, Nevada, or Inyo County, eastern California, but more precise details of geographic and stratigraphic provenance for this specimen are unknown.
Discussion. Lochmanolenellus trapezoidalis is easily distinguished from Lo. primus and Lo. pentagonalis by its trapezoidal rather than pentagonal cephalic outline (Figs 9.1, 9.2, 18.8), and by its more anteriorly positioned genal spine bases (Fig. 9.3). Differences between Lo. trapezoidalis and Lo. subquadratus are listed in the discussion of the latter species. In many respects, Lo. trapezoidalis represents the most extreme morphology known within the genus: the ocular lobes are proportionally shorter than in any other species, and no other species exceeds Lo. trapezoidalis in terms of anterior advancement and inward displacement of the genal spine bases.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Olenellina |
|
Family |
|
|
Genus |