
Haploniscus unicornis Menzies, 1956
|
publication ID |
https://doi.org/ 10.5281/zenodo.201727 |
|
DOI |
https://doi.org/10.5281/zenodo.6185449 |
|
persistent identifier |
https://treatment.plazi.org/id/B01F87C4-FFE4-A23B-FCF5-F8B338B70D9C |
|
treatment provided by |
Plazi |
|
scientific name |
Haploniscus unicornis Menzies, 1956 |
| status |
|
Haploniscus unicornis Menzies, 1956 View in CoL complex
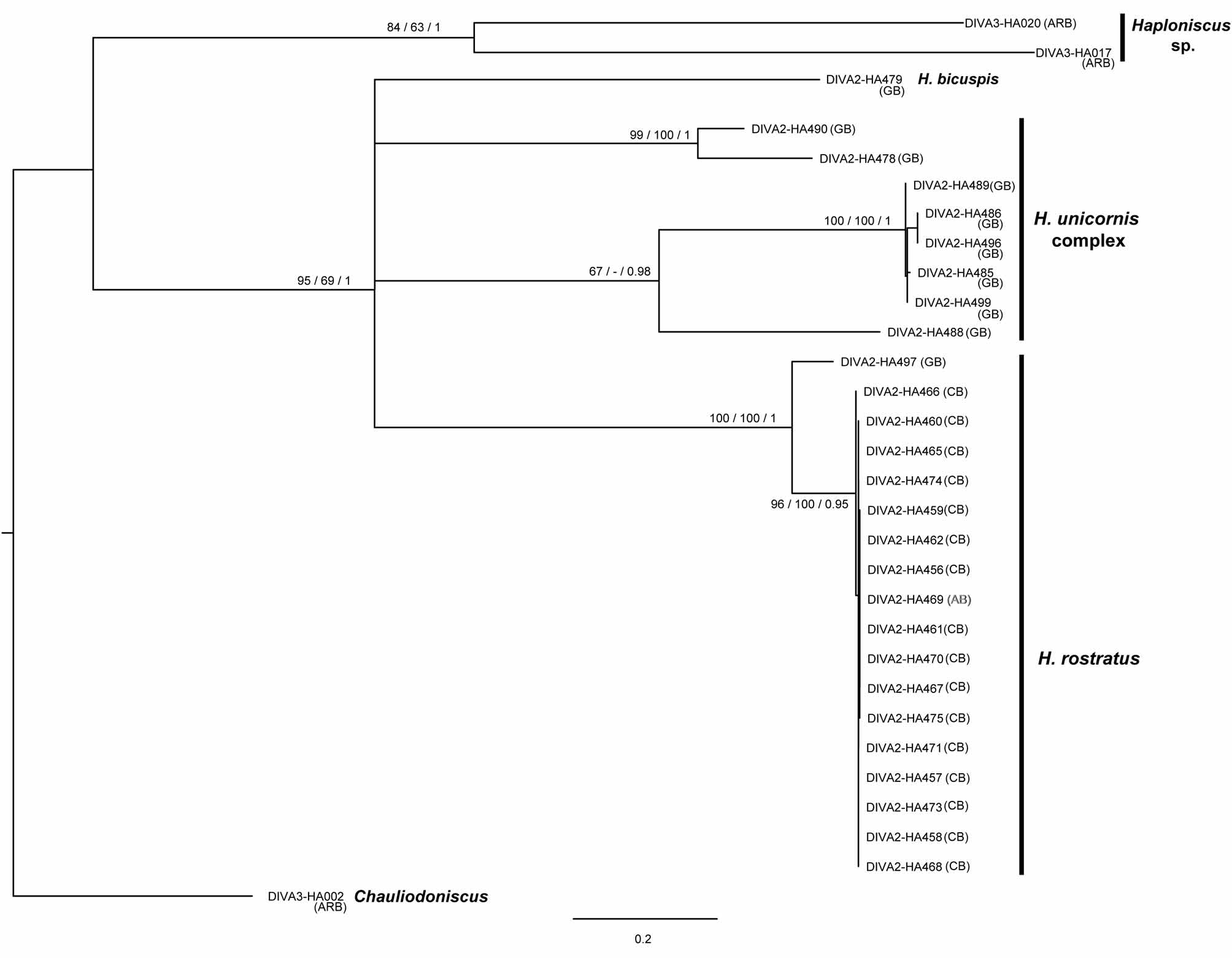
Brökeland (2010b) described four new species belonging to the H. unicornis complex, i.e. H. bihastatus Brökeland, 2010 , H. monoceros Brökeland, 2010 , H. machairis Brökeland, 2010 , and H. angolensis Brökeland, 2010 . All species were described based on male characters. Brökeland (2010b) stated that it was difficult or impossible to assign females, juveniles and manca stages to one of the six species belonging to the complex by morphological traits. In case of the H. unicornis complex we have only sequences of female specimens available, which thus cannot be assigned to one of the species mentioned above, neither using morphological nor genetic characters. The type specimen of H. unicornis was collected from the Puerto Rico trench in the western Atlantic while the specimens analyzed by Brökeland (2010b) were sampled in the Guinea and Angola Basins in the southeastern Atlantic. In contrast to H. rostratus , H. unicornis could not be recorded from the Cape Basin south of the Walvis Ridge. Thus, its distribution is hypothesized to be limited by the Walvis Ridge, in contrast to H. rostratus (see Brökeland 2010a and results above).
Of the six species in the H. unicornis complex, H. monoceros , H. bihastatus and H. machairis were described (based on males) to occur sympatrically in the Guinea Basin (Stations 89 and 90 of the DIVA2-cruise; Brökeland 2010b). In the phylogenetic tree ( Fig. 1 View FIGURE 1 ) the female specimens form no single monophyletic group but cluster in four distinct groups (1: HA490; 2: HA478; 3: HA485, HA486, HA489, HA496, HA499; 4: DIVA2-HA488) separated by 9% in case of group 1 and 2, otherwise by 18–20% sequence divergence (uncorrected p -distance) each. Similarly high pairwise sequence divergence values are found between members of these H. unicornis groups and other distinct species ( H. bicuspis , H. rostratus ). The high between-group divergence is contrasted by a withingroup variability of below 1.8%. The outgroup, a specimen from the haploniscid genus Chauliodoniscus , is separated from the ingroup taxa by 25–28%. A distance of 23.2–24.5% separates Haploniscus sp. from the H. unicornis complex, H. rostratus and H. bicuspis . Comparable values for intra- and interspecific divergences are found also for other asellote isopods ( Osborn 2009; see also supplement 1). This comparison further supports that the H. unicornis groups found in our analysis ( Fig. 1 View FIGURE 1 ) represent different species within the H. unicornis complex and may very likely represent three of the four newly described species described by Brökeland (2010).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Asellota |
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Asellota |
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Asellota |
|
Family |
|
|
Genus |