
Tyrannoseira Bellini & Zeppelini, 2011
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4586.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:F83F9DE6-5DD1-4FD2-8C2E-C01A7F49E870 |
|
persistent identifier |
https://treatment.plazi.org/id/B01387BB-5270-FF8F-61C6-C436FC814390 |
|
treatment provided by |
Plazi |
|
scientific name |
Tyrannoseira Bellini & Zeppelini, 2011 |
| status |
|
Genus Tyrannoseira Bellini & Zeppelini, 2011
Type species. Seira raptora Zeppelini & Bellini, 2006
Diagnosis of genus. Head dorsal macrochaetotaxy lacking M2, M 4i, S1, S4, S5, Ps2 and Pa4 mac ( Figs 3A, E View FIGURE 3 , 4A View FIGURE 4 , 8C View FIGURE 8 ). Four labral papillae, inner papillae bifurcate distally ( Figs 4B View FIGURE 4 , 8E View FIGURE 8 ). Basomedian labial field with chaetae r reduced ( Figs 4A and C View FIGURE 4 , 9C View FIGURE 9 ). Th II with 3–4 medio-median mac (m1, m 1i, m2, m 2i) and 2–3 medio-lateral mac (m4, m 4i, m4p); groups PmA–PmC with 4 –5, 2–3 and 3–5 mac, respectively ( Figs 10A View FIGURE 10 , 15A View FIGURE 15 , 30A View FIGURE 30 ). Th III with at least 7 central mac (a2, a4–5, p1–3), a6 mac or mic ( Figs 10B View FIGURE 10 , 15B View FIGURE 15 , 20B View FIGURE 20 ). Abd I only with mic ( Fig. 10C View FIGURE 10 ). Abd II with 3 central mac (a2, m3, m3e), m3ep absent ( Fig. 10D View FIGURE 10 ). Abd III with 1 central (m3) and 3 lateral mac (pm6, p6 and p6e exclusively) lateral to the bothriotricha, am6 as mic ( Fig. 10E View FIGURE 10 ). Abd IV with at least 8 central mac (A3, A5, B3–6, C1, T1), A3a, B1 and B2 as mac or mic ( Figs 11A View FIGURE 11 , 16A View FIGURE 16 , 21A View FIGURE 21 , 31A View FIGURE 31 ). Leg I of males internally with 1 finely ciliate trochanteral spine, femur strongly enlarged and with one subdistal group of spines heavily ciliate, tibiotarsus I armed with an inner row of finely ciliate proximal spines ( Figs 5 View FIGURE 5 A–C, 12 View FIGURE 12 E–F). Collophore anteriorly with proximal smooth spine-like chaetae, posteriorly with distal spines ( Figs 6B View FIGURE 6 , 13A View FIGURE 13 ).
Remarks. The distinguishing characters of Tyrannoseira species are the fore legs of adult males, with enlarged femora each carrying one subdistal group of inner spines. Both characters are unique for the genus while others presented in the diagnosis are present on single species or some are found in combination in species of Seira s. lat. Although some species of the domestica -group can also have superficially similar dimorphic legs on males (but with femoral spines only in one longitudinal row), their dorsal macrochaetotaxy is clearly different, suggesting they belong to different lineages of Seirinae ( Bellini & Zeppelini 2011 , Cipola et al. 2018b).
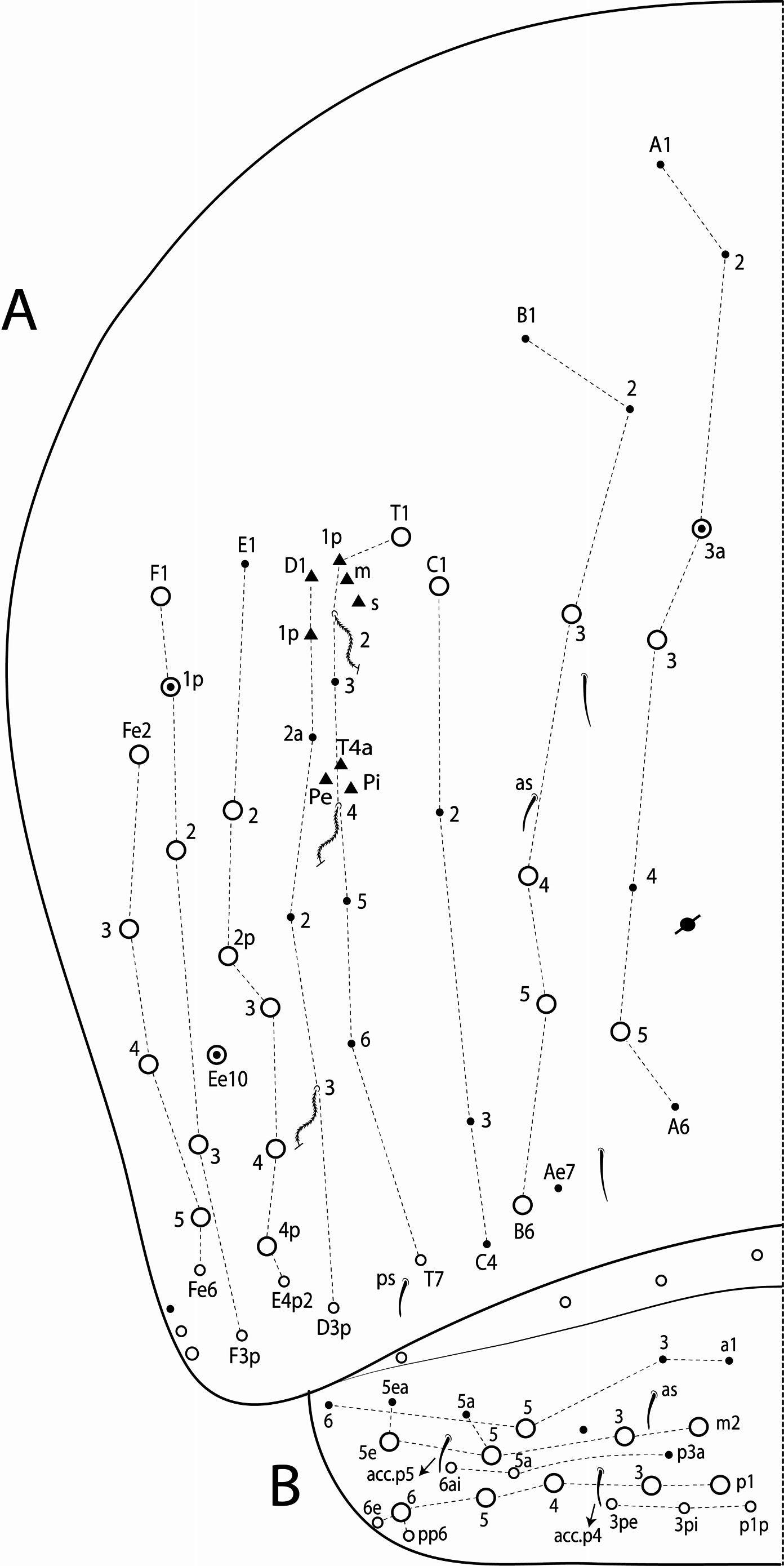
Morphological characters of Tyrannoseira . The following characters are shared by all five Tyrannoseira species and are not repeated in the descriptions.
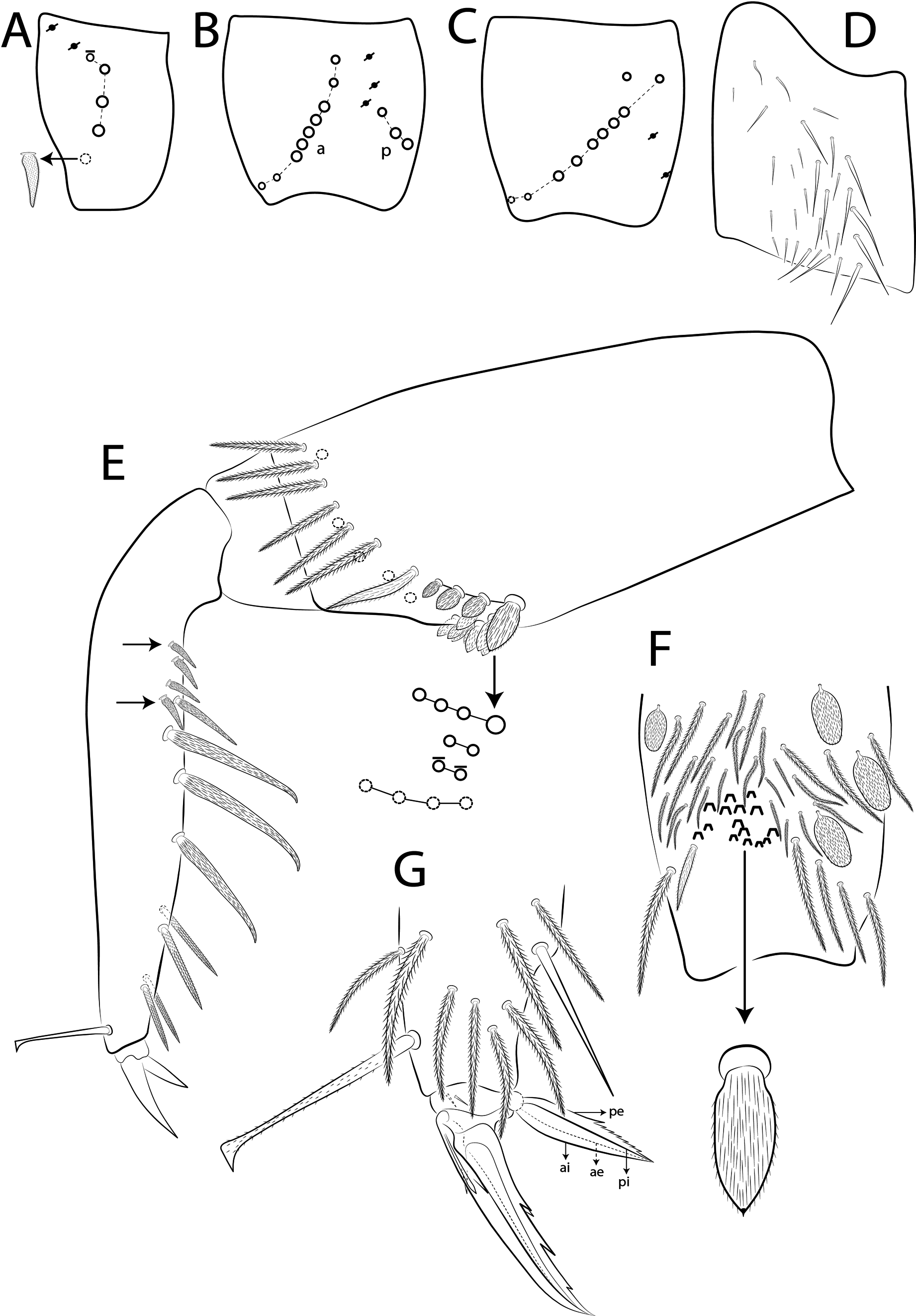
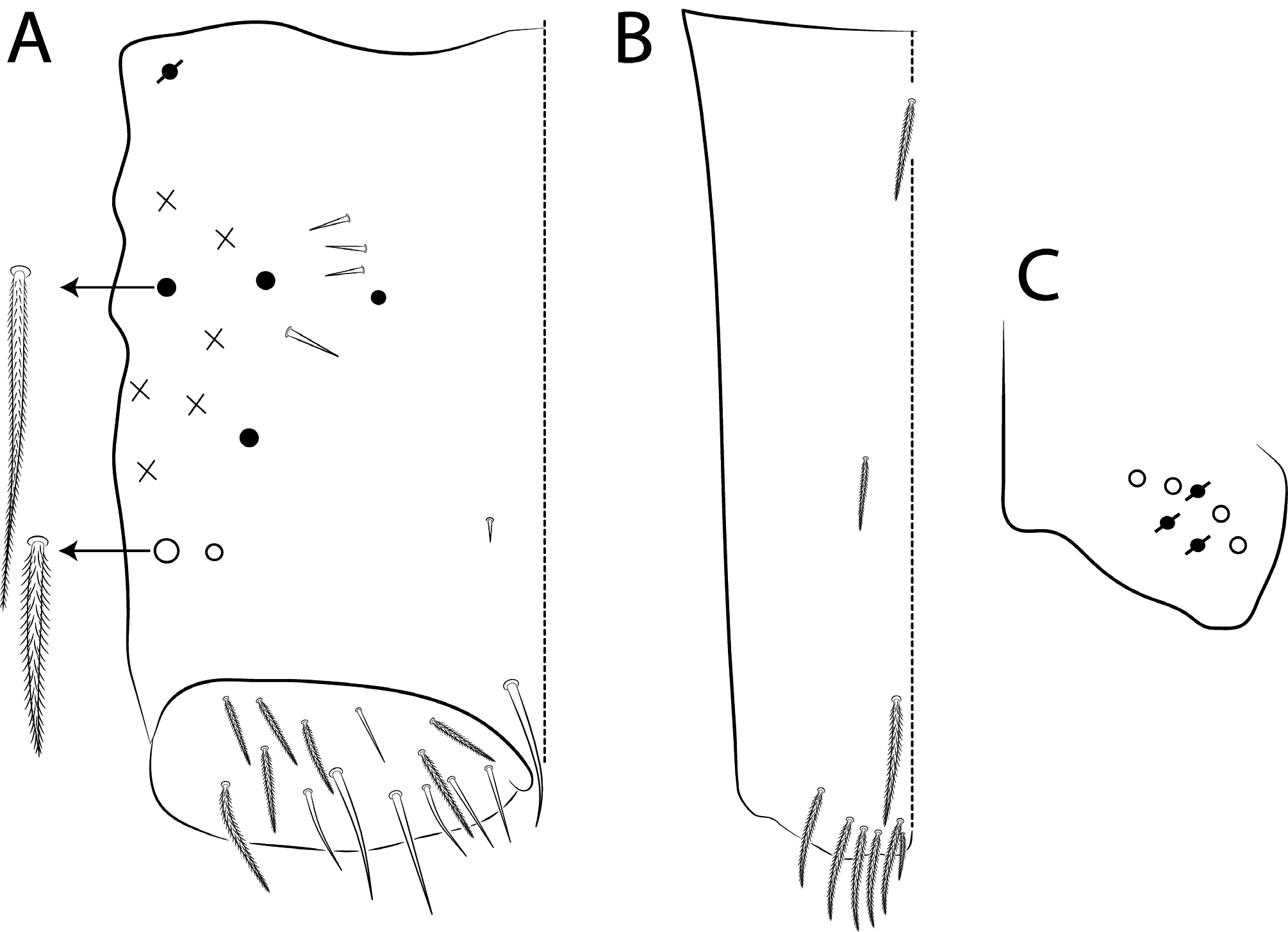
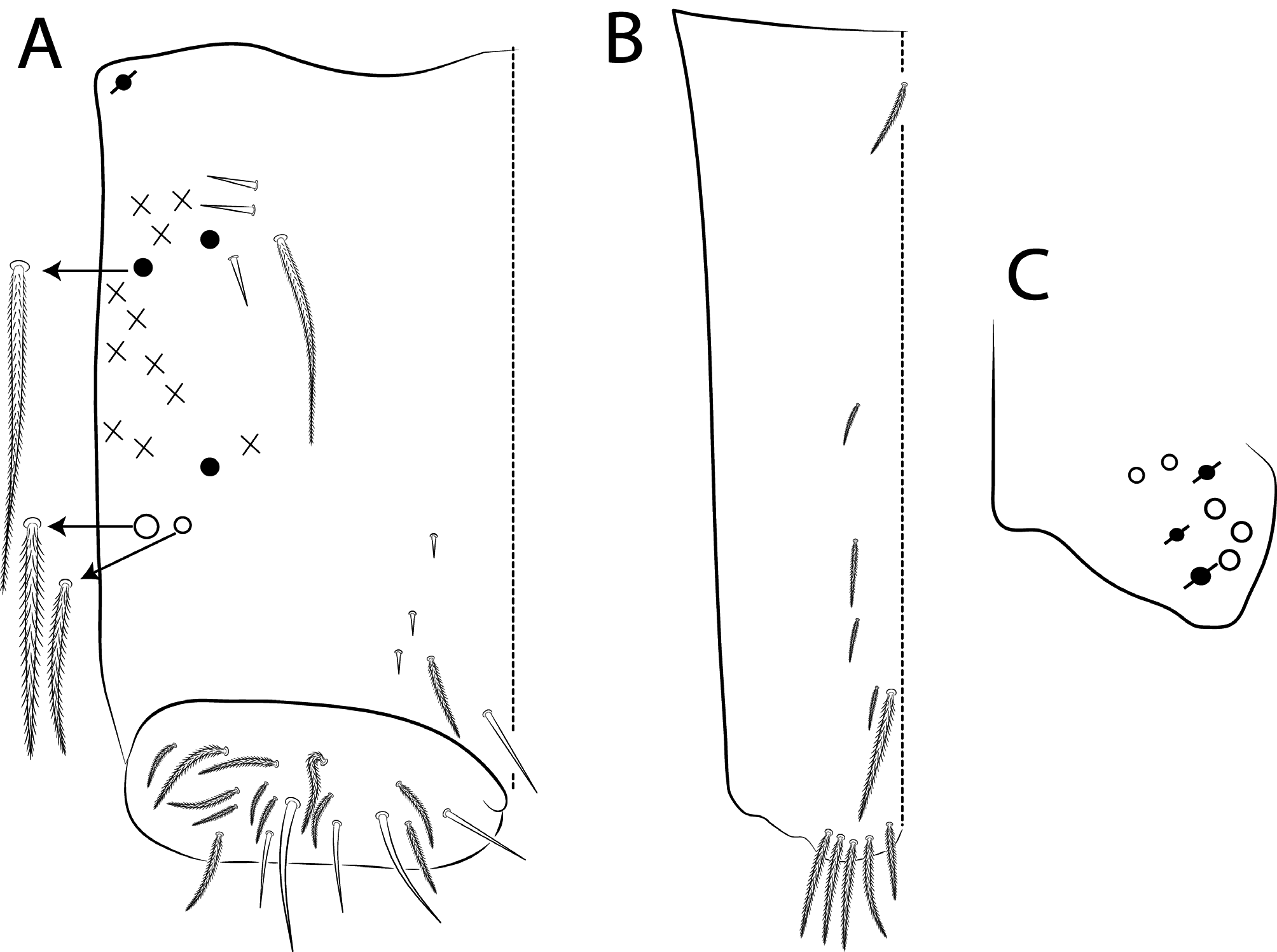
Dorsal chaetae and structure with bothriotricha ( Figs 3A, E View FIGURE 3 , 7A View FIGURE 7 , 8C View FIGURE 8 , 10 View FIGURE 10 D–E, 11A View FIGURE 11 ) as densely ciliate but elongated cilia, short on head (1 subantennal and 1 post-ocellar) and elongated on abdomen; Abd II–IV formula with 2 (a5, m2), 3 (a5, m2, m5) and 3 (T2, T4, D3) bothriotricha. Bothriotrichal accessory chaeta ( Figs 2I View FIGURE 2 , 7A View FIGURE 7 , 10 View FIGURE 10 D–E, 11 View FIGURE 11 A), heavily ciliate on distal two thirds or more, with fringed aspect and heavily ciliated on Abd II–IV. Mac ( Figs 2B View FIGURE 2 , 3A View FIGURE 3 , 7A View FIGURE 7 ). Finely ciliate and apically truncate-shaped mac present on dorsal head and Th II to Abd V. Mes ( Figs 1 View FIGURE 1 A–C, 7A View FIGURE 7 ) heavily ciliate and apically acuminate, present on body, generally in Abd IV. Mic ( Figs 2 View FIGURE 2 D– J, 7A View FIGURE 7 , 8C View FIGURE 8 , 10–11 View FIGURE 10 View FIGURE 11 ) smooth, apically acuminate and of similar lengths on different regions of the body, but with different forms, simple, with 1 median ciliation, with 4 unilateral ciliations, or with 7 or more bilateral ciliations, present on head and AMP series of Th II to Abd V. Ordinary sens ( Figs 2C View FIGURE 2 , J–K, 7A View FIGURE 7 , 10–11 View FIGURE 10 View FIGURE 11 ), smooth and apically rounded of similar lengths, present on Th II–III (al), Abd II–III (as, acc.p6), Abd IV (as, ps) and Abd V (as, acc.p4, acc.p5). Th II–Abd V sens formula 1, 1 | 0, 2, 2, +, 3. Psp ( Figs 2L View FIGURE 2 , 7A View FIGURE 7 , 10–11 View FIGURE 10 View FIGURE 11 ), as small, circular, smooth concavity with a narrow opening, present centrally on Th II to Abd IV and on coxae and manubrial plates. Specialised microchaetae – ms ( Figs 2C, H View FIGURE 2 , 7A View FIGURE 7 , 10–11 View FIGURE 10 View FIGURE 11 ) as small, smooth and distally striate chaetae with rounded apex present on Th II, Abd I and III. Th II–Abd V, ms formula 1, 0| 1, 0, 1, 0, 0.
Scales ( Figs 2A View FIGURE 2 , 3A View FIGURE 3 , 4A View FIGURE 4 , 5 View FIGURE 5 A–B, 6 View FIGURE 6 F–G, 7 View FIGURE 7 A) heavily ciliate with short interrupted cilia, generally oval or elongated and apically rounded (rarely truncate, pointed or irregular). The cilia are toothpick-shaped and abruptly acuminate at apex. Scales present on Ant I–III, both sides of head, dorsal and lateral trunk, all legs (except pretarsus), anterior collophore and furcula ventrally to lateral margin.
Antennal segments with at least 8 types of chaetae, as shown in Fig. 7B; a–f View FIGURE 7 as sens, g–h as ciliate chaetae: type a pair of rods or clubs apical organ present laterally on Ant III; type b s mooth, elongated and slightly apically acuminate, present on Ant II–IV ( Fig. 3D View FIGURE 3 ;, type c s mooth, somewhat short and finger-shaped, present densely on Ant I–IV ( Figs 3 View FIGURE 3 C–D); type d s mooth, short and finger-shaped, present densely on Ant I–IV ( Figs 3 View FIGURE 3 C–D); type e three guard sens, smooth and spiny-like surrounding apical organ of Ant III ( Fig. 3D View FIGURE 3 ); type f. striated and rounded, subapical modified sens of Ant III ( Fig. 3D View FIGURE 3 ).; type g weakly ciliate, present on Ant I–IV ( Fig. 3D View FIGURE 3 ); type h heavily ciliate, present on Ant I–IV ( Fig. 3D View FIGURE 3 ).
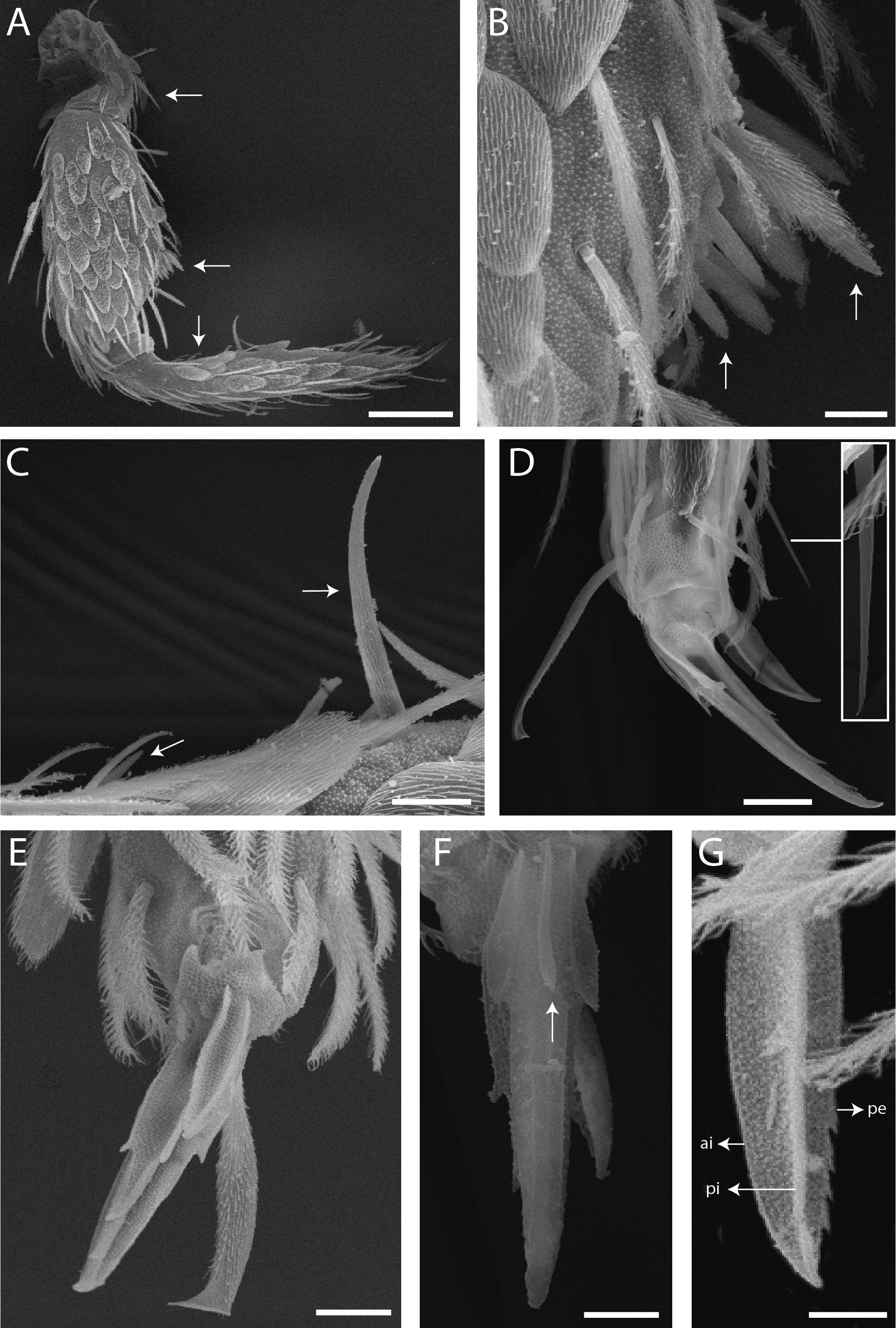
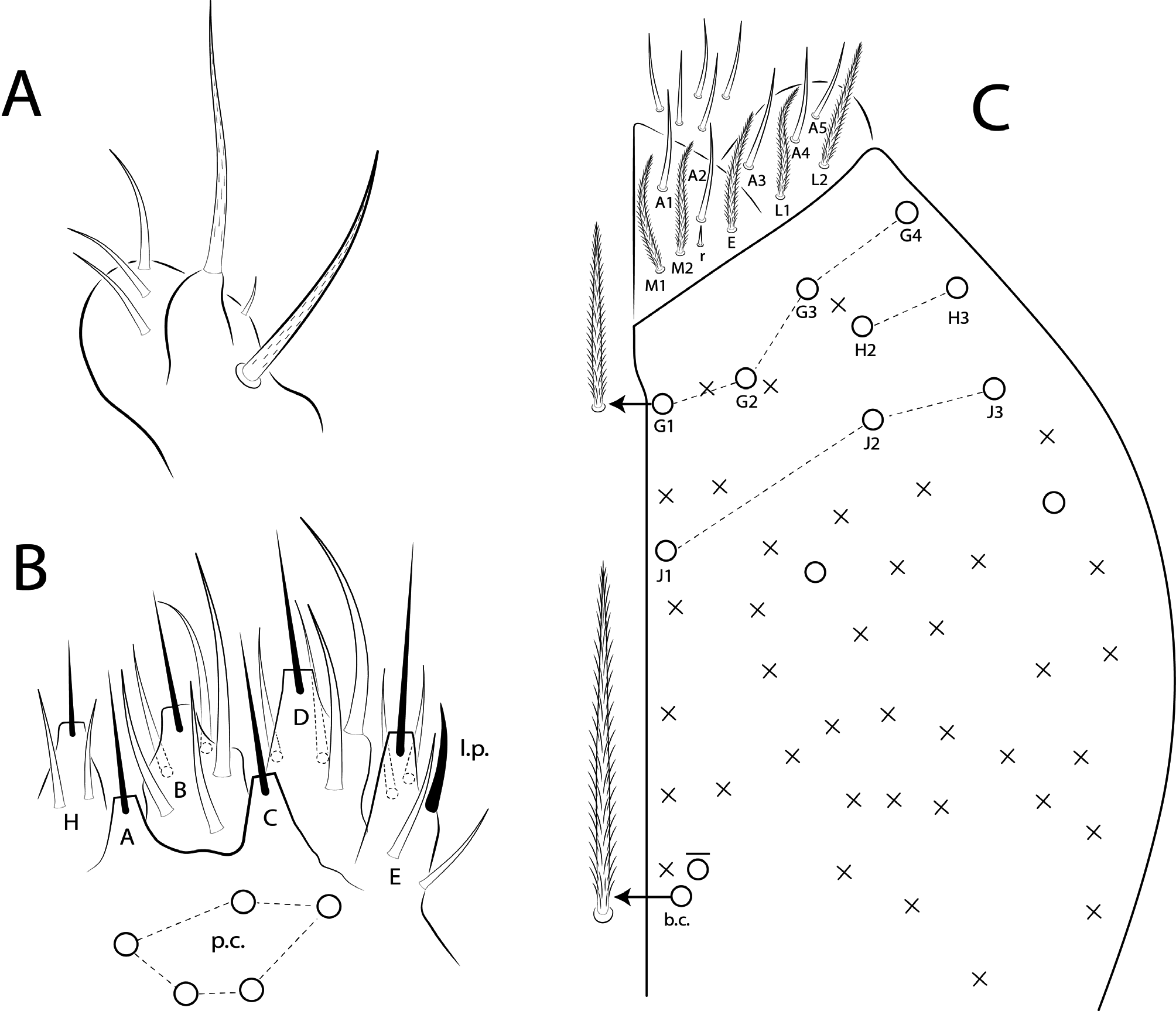
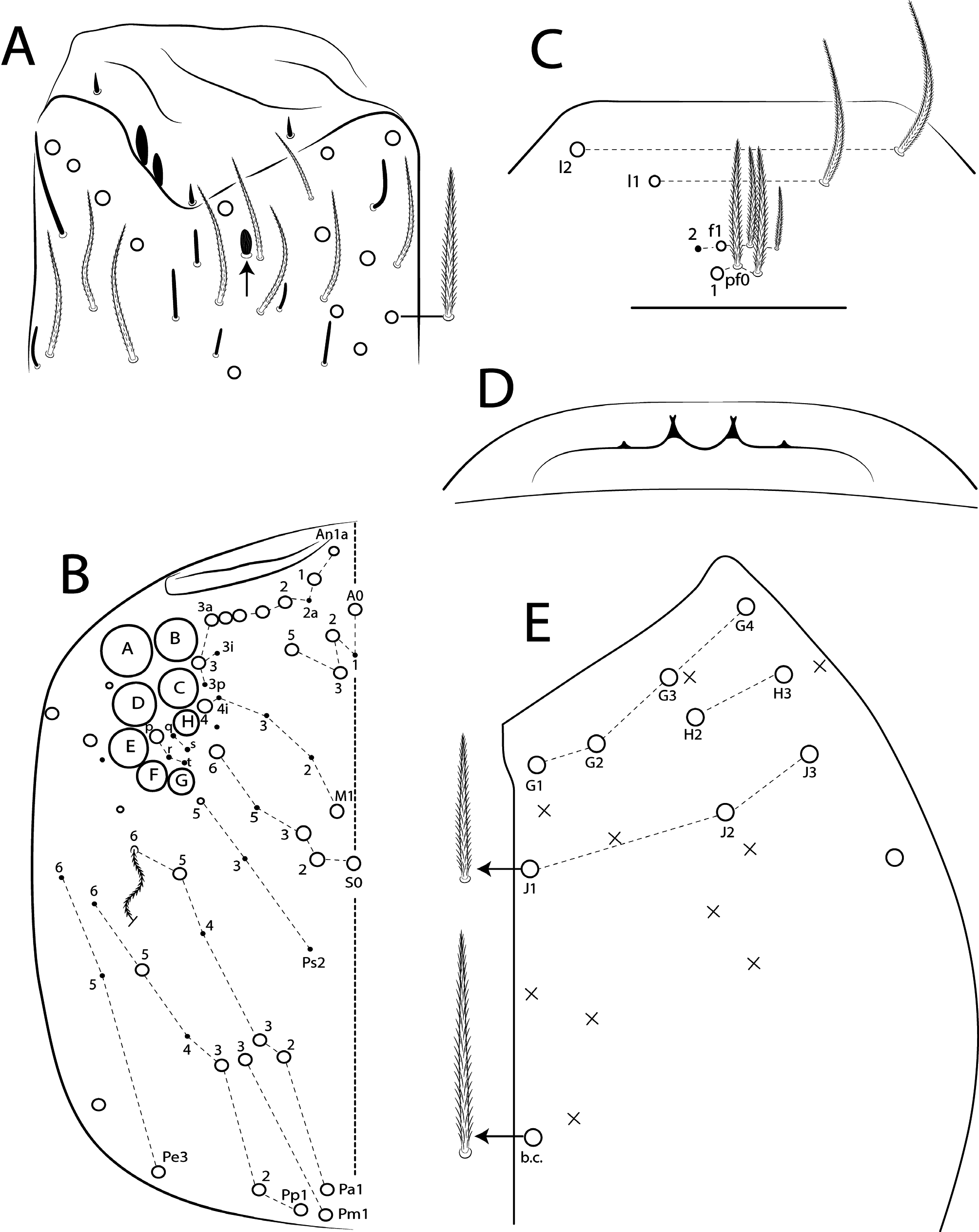
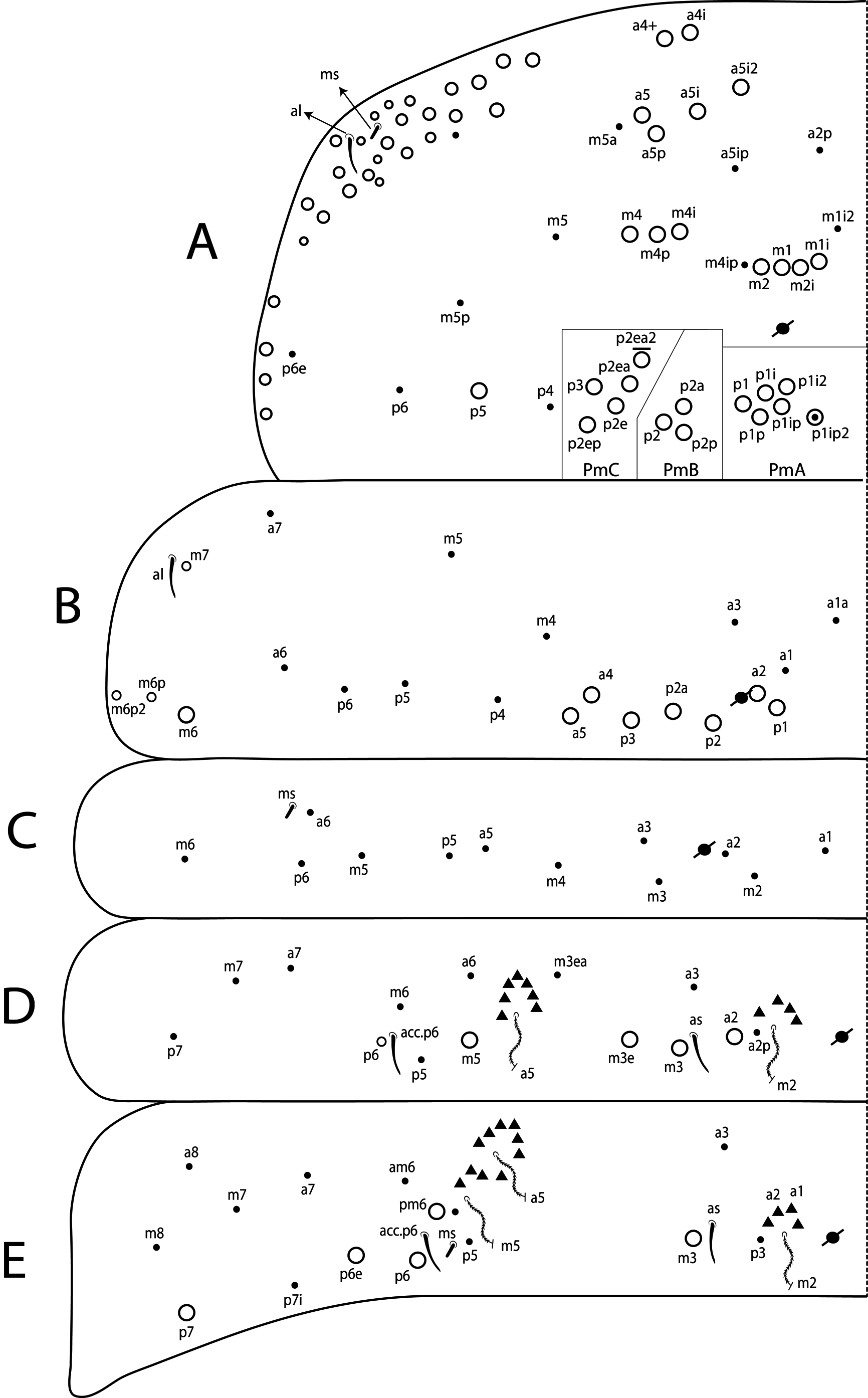
Antennae shorter than body length; Ant IV longer or subequal to Ant II–III, Ant I shorter ( Fig. 1 View FIGURE 1 ); Ant IV not annulated but slightly ringed; distal, bilobed apical bulb present ( Figs 3 View FIGURE 3 B–C, 8 View FIGURE 8 A). Eyes oval, 8+8 per side and with 5 interocular chaetae (q, s, p, r, t) ( Figs 3A View FIGURE 3 , 8C View FIGURE 8 ); dorsal macrochaetotaxy ( Fig. 8C View FIGURE 8 ) with 6–9 ‘An’ (An1a–3), 4 ‘A’ (A0, A2–3, A5), 2 ‘M’ (M1, M4), 4 ‘S’ (S0, S2–3, S6), 4 ‘Pa’ (Pa1–3, Pa5), 2 ‘Pm’ (Pm1, Pm3), 4 ‘Pp’ (Pp1– 3, Pp5) and at least 1 ‘Pe’ (Pe3); microchaetotaxy with 1 ‘An’ (An2a), 1–2 ‘A’ (A1, A4), 3 ‘M’ (M2– 4i), 1‘S’ (S5), 2 ‘Ps’ (Ps2– 3), 1 ‘Pa’ (Pa4), 2 ‘Pp’ (Pp4, Pp6) and 1–2 ‘Pe’ (Pe5–6); chaetae S1 and S4 absent. Four prelabral ciliate chaetae of subequal length; labral formula with 4 (a1–2), 5 (m0–2), 5 (p0–2) smooth chaetae, a1 apically rounded, p0–1 larger, others subequal ( Figs 4A View FIGURE 4 , 8D View FIGURE 8 ). Labral papillae conical and apically rounded, inner papillae bifurcate distally, outer papillae simple ( Figs 4 View FIGURE 4 A–B, 8 View FIGURE 8 E, 14 View FIGURE 14 D). Labial palp with five main papillae (A–E) plus one hypostomal papilla (H) with 0, 5, 0, 4, 4, 2 guard appendages respectively; lateral process (l.p.) apically acuminate and surpassing the base of apical appendage ( Figs 4D View FIGURE 4 , 9B View FIGURE 9 ). Labium with five smooth proximal chaetae (p.c.) ( Figs 4D View FIGURE 4 , 9B View FIGURE 9 ). Basolateral and basomedian labial fields with chaetae a1–5 smooth, M1–2, E, L1–2 ciliate, r spine-like, reduced and smooth ( Figs 4A and C View FIGURE 4 , 9C View FIGURE 9 ). Maxillary palp with apical appendage (a.a.) and basal chaeta (b.c.) slightly ciliate and subequal in size; sublobal plate internally with 3 main, smooth appendages, externally with 1 minute, smooth appendage ( Figs 4A View FIGURE 4 , 9A View FIGURE 9 ).
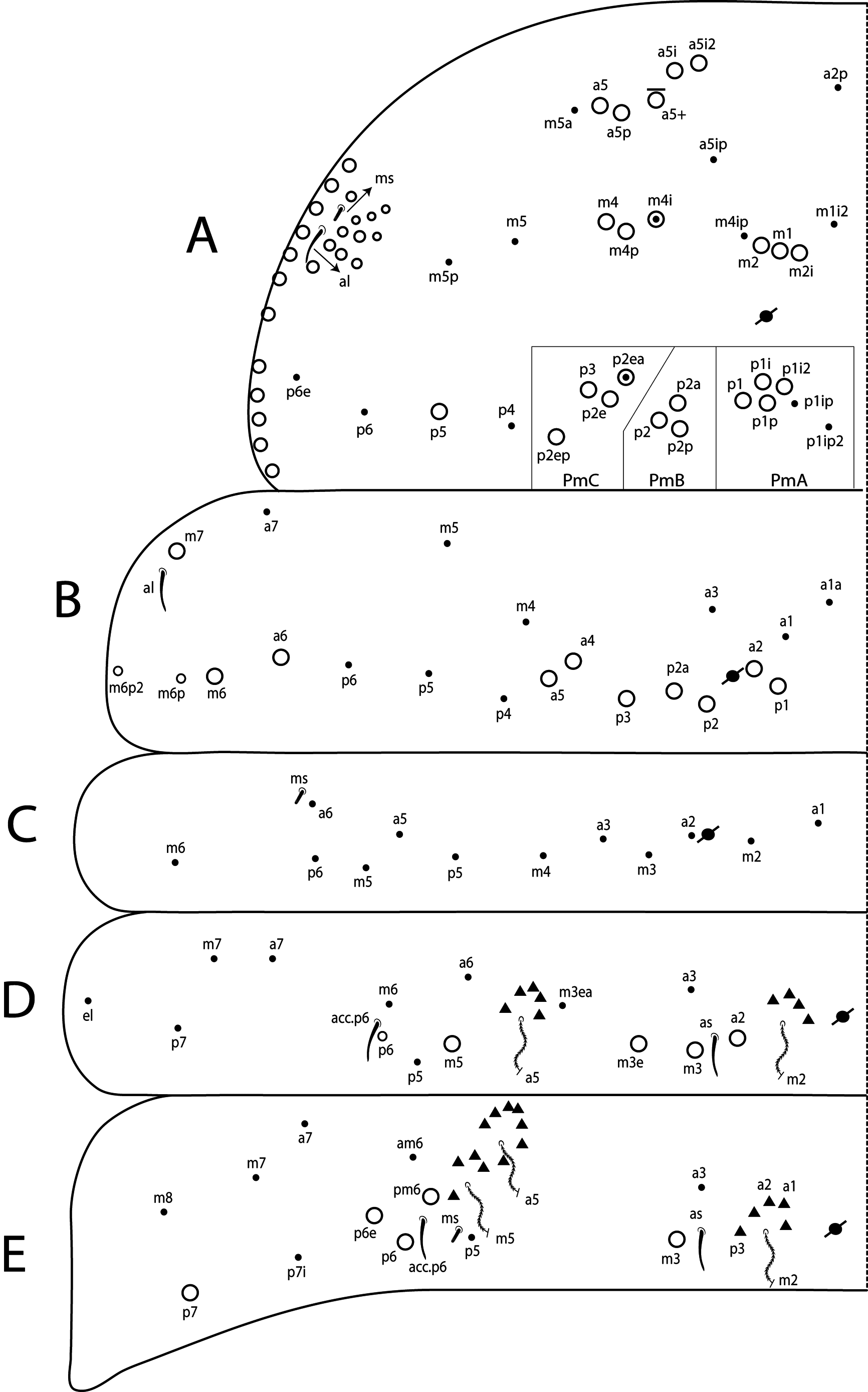
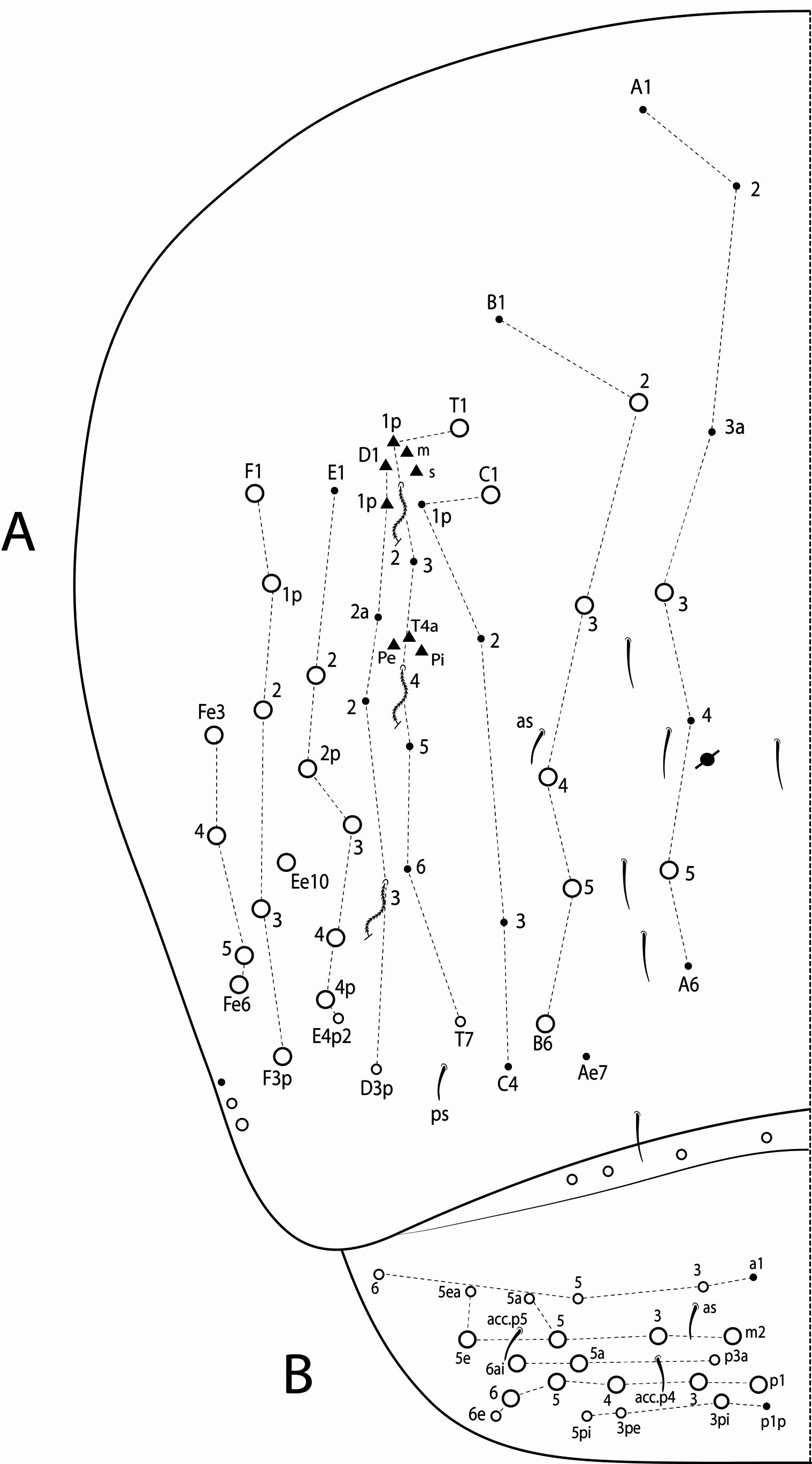
Thoracic chaetotaxy ( Figs 10 View FIGURE 10 A–B, 15 View FIGURE 15 A–B, 20 View FIGURE 20 A–B, 25 View FIGURE 25 A–B, 30 View FIGURE 30 A–B). Th II with 21–28 main mac (a 4i, a4+, a5ip–5p, m1– 1i, m2– 2i, m 4i –4p, p1ip2–1p, p2a–2p, p2ea2–2ep) and 10 main mic (a2p, a5ip, m 1i 2, m4ip, m5a– 5p, p4, p6–6e), m 1i present or absent, m4p rarely as mic, p1ip2 rarely as mac, p 1i 2, p1ip, p2p, p2ea and p2ea2 (present or absent) as mac or mic. Th III with 8–11 mac (a2, a4–6, m6–6p, m7, p1, p2–2a, p3) and 8–10 mic (a1a– 1, a3, a6–7, m4–5, p 1i, p4–6), a6 as mac or mic, a3 rarely as mac and p 1i present or absent.
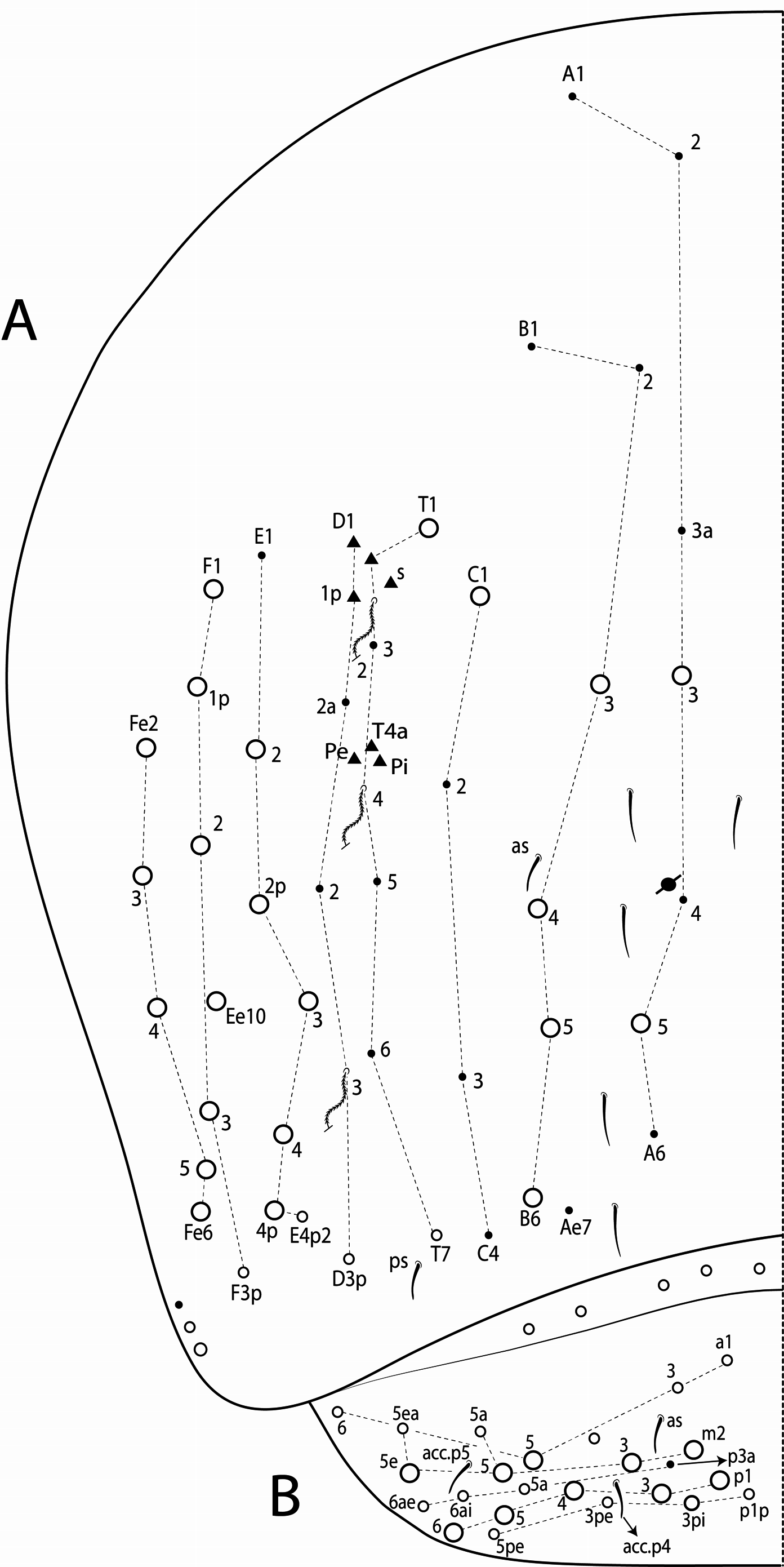
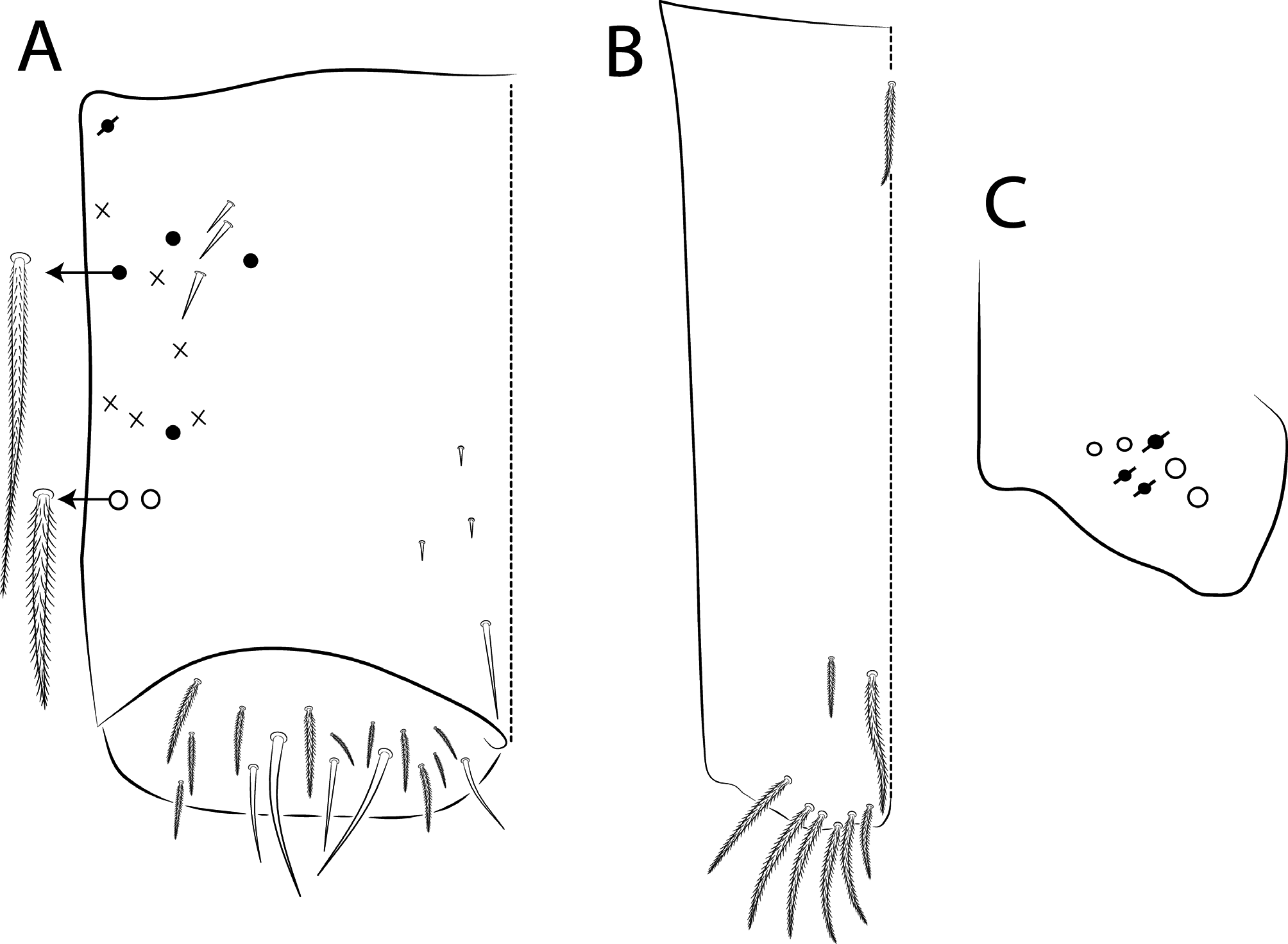
Abdominal chaetotaxy ( Figs 10 View FIGURE 10 C–E, 11 View FIGURE 11 A–B, 15 View FIGURE 15 C–E, 16 View FIGURE 16 A–B, 20 View FIGURE 20 C–E, 21 View FIGURE 21 A–B, 25 View FIGURE 25 C–E, 26 View FIGURE 26 A–B, 30 View FIGURE 30 C–E, 31 View FIGURE 31 A– B). Abd I with 12 mic (a1–3, a5–6, m2–6, p5–6). Abd II with 4 mac (a2, m3, m3e, m5), 1 mes (p6), 8–10 mic (a2p, a3, a6–7, m3ea, m6–7, p5, p7, el), and bothriotricha a5 and m2 with 5–7 and 3–4 accessory chaetae, respectively; chaeta a2p rarely present, p7 and el rarely as mes. Abd III with 5 mac (m3, pm6, p6, p6e, p7), 6–8 mic (a3, a7, a8, am6, m7, m8, p5, p 7i), bothriotrichum m2 with 4–5 accessory chaetae (a1–2 and 2 unnamed, p3 accessory or mic) and bothriotricha a5 and m5 with 10–11 accessory chaetae between them (em, emp and 8 unnamed, c3 accessory or mic), a8 and m8 present or absent, p 7i mes or mic rarely absent, p8 rarely present, p3 as accessory chaeta or mic, present or absent. Abd IV with 21–29 mac (A3a–3, A5, B1–6, C1, T1, T7, D3p, E2–4p, Ee10, F1–3p, Fe2–6), about 15 mic (A1–2, A4, A6, Ae7, C1p–4, T3, T5–6, D2a–2, E1), bothriotricha T2 and T4 with 4–5 (s, m, T1p, D1–1p) and 3 (T4a, Pi, Pe) accessory chaetae, respectively, A3a, B1, B2, T6, Ee10 and F1p as mac or mic, D3p and F3p as mes or mic, C1p rarely present, m present or absent; posteriorly with 4–5 mes. Abd V with 9–14 mac (a5, m2, m3, m5, m5e, p1, p3–6e, p3pi, p5a, p6ai) and 10–16 mes or mic (a1, a3, a6, m5a, m5ea, p3a–6ae, p1p–5pe, pp6), a5 as mac, mes or mic, p5a, p6ai and p3pe as mac or mes, p4a, p5ae, p5pi, p5pe, p6e and pp6 present or absent.
Legs with subcoxa I having one row of ciliate chaetae and 2 psp; subcoxa II with two rows (anterior and posterior) of ciliate chaetae and 3 psp; subcoxa III with one row of ciliate chaetae and 2 (rarely 3) psp ( Figs 12 View FIGURE 12 A– C). Leg I coxa in males with 1 subdistal, finely ciliate inner spine, apically pointed ( Fig. 12A View FIGURE 12 ). Trochanter with 1 inner, club-shaped spine, finely ciliate ( Fig. 5A View FIGURE 5 ). Femur strongly enlarged on outer and inner faces and with one subdistal group of heavily ciliate inner spines, varying in thickness and apically pointed ( Figs 5 View FIGURE 5 A–B). Tibiotarsus armed by a single inner row of finely ciliate, apically rounded spines (proximal shorter, median larger and apically curved), also with heavily ciliate and apically acuminate distal spine-like mac ( Figs 5A, C View FIGURE 5 , 12E View FIGURE 12 ); outer face distally with 1 finely ciliate, apically capitate, tenent hair, inner face of tibiotarsus III with one weakly grooves smooth chaetae on distal two thirds ( Figs 5 View FIGURE 5 D–E). Pretarsus with one minute anterior and one posterior smooth chaetae ( Fig. 12G View FIGURE 12 ). Unguis inner face with 2 paired basal teeth, 1 unpaired median tooth, and 1 unpaired apical tooth; outer face with 2 paired lateral teeth and one unpaired apically pointed basomedian lamella ( Figs 5 View FIGURE 5 D–F). Unguiculus with 4 acuminate lamellae, pe lamella serrated on distal half and with a small median tooth, other lamellae smooth (ai, ae, pi) ( Figs 5E, G View FIGURE 5 , 15G View FIGURE 15 ). Collophore ( Figs 6 View FIGURE 6 A–B, 13A View FIGURE 13 , 28A View FIGURE 28 ) with 9–10 chaetae on anterior face, proximal region with 1 psp, 3–4 reduced spine-like smooth chaetae and at least 4 larger, ciliate, apically acuminate chaetae, plus 1–2 ciliate mac distally of equal or differing lengths; posterior face with 2–5 distal chaetae, 1–3 smooth spines, 1 ciliate chaeta present or absent plus 1 smooth chaeta; lateral flap with distal smooth chaetae and proximal ciliate chaetae. Tenaculum with 4 teeth on each ramus; corpus with one weakly ciliate chaeta. Genital plate circinate in males, with 15 circumgenital smooth chaetae (1 superior unpaired) and 3+3 small eugenital smooth chaetae ( Figs 6C View FIGURE 6 , 13B View FIGURE 13 ). Female with two pairs (superior and inferior) of small smooth chaetae, without other modifications. Furcula ( Figs 1 View FIGURE 1 , 6D View FIGURE 6 ) subequal to length of abdomen. Manubrium shorter than dens and with ciliate chaetae; ventral face with ciliate chaetal formula specific to each species ( Figs 13C View FIGURE 13 , 18B View FIGURE 18 , 23B View FIGURE 23 , 28B View FIGURE 28 , 33B View FIGURE 33 ); dorsal face densely covered with ciliate chaetae of different lengths, manubrial plate distally with up to 5 ciliate chaetae and 3 psp. Dens dorsally crenulate with numerous ciliate chaetae, lacking other modifications such as spines or tubercles ( Fig. 6E View FIGURE 6 ). Mucro dorsally falcate, without basal spine ( Fig. 6E View FIGURE 6 ).
Remarks. Corrections to the character descriptions include the apical bulb of Ant IV which was originally described as absent in T. diabolica and unilobed in other species, but it is bilobed in all species ( Figs 3C View FIGURE 3 , 8A View FIGURE 8 ). From lateral view the apical bulb of Ant IV can be misinterpreted as having a single lobe (e.g. Zeppelini & Lima 2012: 38, fig. 2), because the lobes can overlap, and so they only the dorsal or ventral face be viewed. Other character typical of all Tyrannoseira spp. the chaeta r is spine-like and reduced in basomedian labial field, but it was reported as absent in T. diabolica , ciliate in T. gladiata and smooth in T. raptora . It is clear in the MEV photograph that the r chaeta is reduced and partially obscured by the a2 smooth chaeta. In the original description r chaeta was mistakenly identified as a2 chaeta (see Zeppelini & Bellini 2006: 25, right face of fig.4). Similarly T. diabolica was reported to have 3 inner teeth on the ungues, but it bears 4 teeth as do all other species.
A detailed chaetotaxy for all species, as done here, needed to be described so that they could be confidently compared. On the head there are 4 mac on ‘A’ series (A0, A2, A3, A5), but A5 was originally omitted in the descriptions of T. raptora and T. sex (see Zeppelini & Bellini 2006: 27, fig. 8a; Bellini & Zeppelini 2011: 553, fig. 6), and two A0 was a misinterpretation of the frontal head of T. gladiata (see Zeppelini & Lima 2012: 41, fig. 17)., The postocular mac (Pa5 and Pp5), as well as the five posterior mac (Pa1, Pm1, Pp1, Pp2, Pe3) also dorsally on the head, were partly omitted from the descriptions, except for T. gladiata . On Th II anteriorly, T. raptora may have 1 extra mac (a5+), while T. diabolica , T. gladiata and T. sex have 2 extra mac (a4+ and a 4i). On Abd II there are 3 central mac (a2, m3 and m3e) in all species ( Figs 10D View FIGURE 10 , 15D View FIGURE 15 , 20D View FIGURE 20 , 25D View FIGURE 25 , 30D View FIGURE 30 ), but T. bicolorcornuta was previously reported with 5 and T. gladiata with 2 mac (see Zeppelini & Lima 2012: 41, fig. 17). Laterally on Abd III there are 4 mac typically present (pm6, p6, p6e and p7), while only 2 or 3 lateral mac have been reported for all species except for T. bicolorcornuta ) in this region. As for Abd IV macrochaetotaxy, T. bicolorcornuta and T. diabolica were reported to have 12 and 13 central mac, respectively. However, number of central mac on Abd IV is at most 10 including 8 central mac (A3, A5, B3–6, C1, T1) which is constant for the genus (e.g. Fig. 11A View FIGURE 11 ), and further mac (A3a, B1–2, T7) are constant for specific species only ( Figs 16A View FIGURE 16 , 21A View FIGURE 21 , 26A View FIGURE 26 , 31A View FIGURE 31 ). The B2 mac is constant only in T. bicolorcornuta , T. gladiata and T. sex , while the other mac can have intraspecific variations that are present or absent as A3a in T. diabolica , B 1 in T. sex , and T 7 in T. gladiata . Other variations thatwere not reported in the original descriptions follow. In ‘m’ series of Th II, m 4i mac was observed as present or absent in T. raptora , while m 1i in T. bicolorcornuta can be present or absent as well. On the same segment, in the PmA–PmC group, the mac that may be present or absent are: p1ip 2 in T. sex , p2ea in T. raptora , p 1i 2 and p2p in T. bicolorcornuta , and p2ea 2 in T. gladiata and T. sex ( Figs 10A View FIGURE 10 , 15A View FIGURE 15 , 20A View FIGURE 20 , 25A View FIGURE 25 , 30A View FIGURE 30 ). Note that out of these variations on both segments (Th II and Abd IV), only the latter (p2ea 2 in Th II) is variable in more than one species, and for this reason the remaining variations can help in the species diagnoses.
Other characteristics omitted from the original descriptions that are now useful to separate species are: Ant III modified subapical sens, labral papillae shape, chaetotaxy of clypeus, ventral head, collophore, manubrial ventral formula and manubrial plate (see Table 1 View TABLE 1 ).
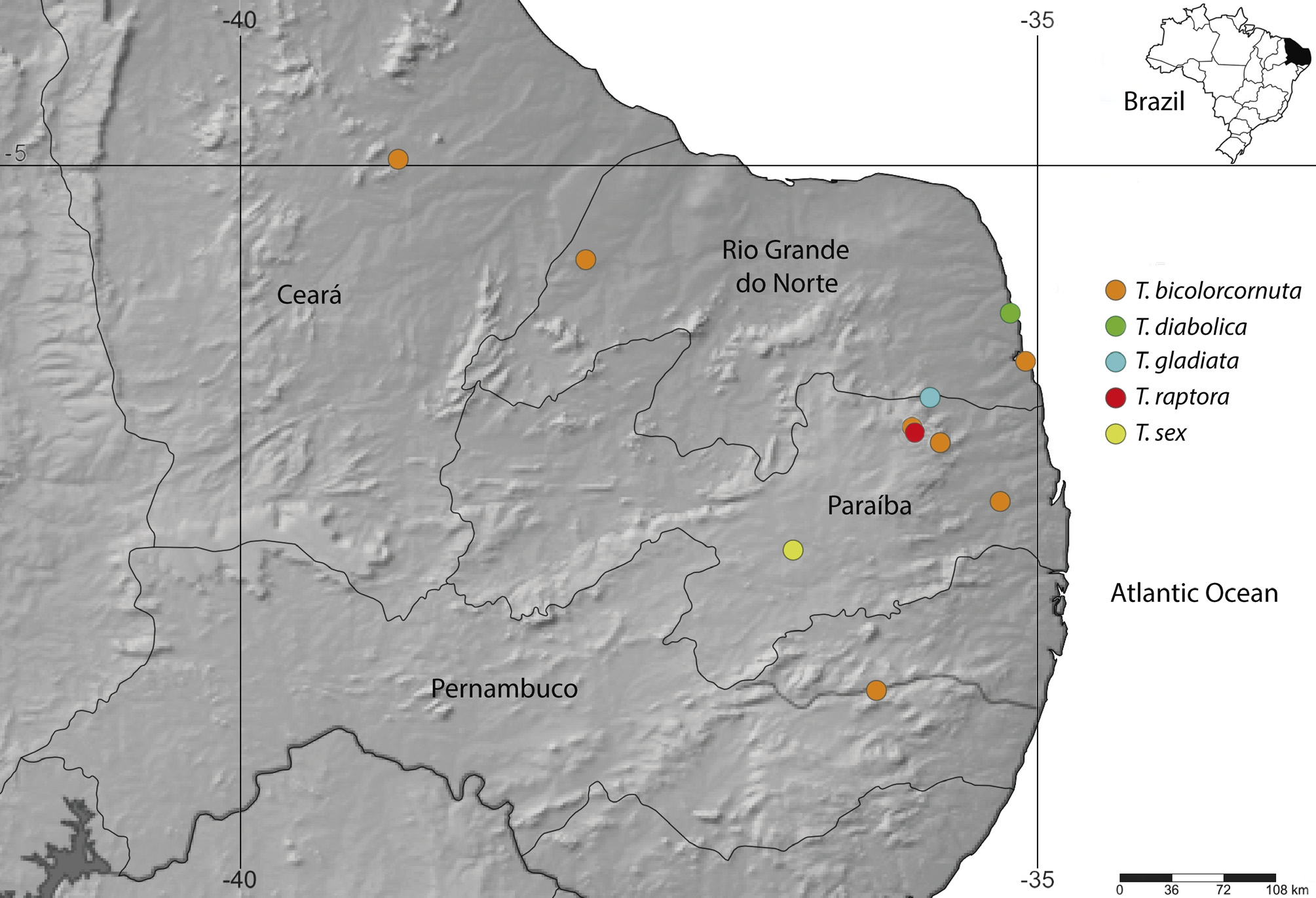
Distribution and habitat of Tyrannoseira . The genus has so far only been found in the northeastern region of Brazil, from Ceará (new record), Paraíba, Pernambuco and Rio Grande do Norte states ( Fig. 34 View FIGURE 34 ), i.e. Good’s biogeographic zone 27 of the Neotropical region ( Good 1974). The regional climate is tropical with a dry season (Aw) and is generally hot semi-arid (BSh), as in northern Atlantic Rainforest (low humidity) and Caatinga (low rainfall) biomes, respectively ( Kottek et al. 2006).
Tyrannoseira is apparently endemic to this region of Brazil, but its preferred microhabitat is still uncertain.
Most species were found living among granitic rocks, even at times of high solar incidence, i.e. above 40°C in the Caatinga phytogeographic domain. Only T. bicolorcornuta (in part) and T. diabolica were found in Atlantic Rainforest. The northern Atlantic Rainforest of Neotropical region suffers from the strong influence of Caatinga, which may be the reason these latter two species occur there.














No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.