
Batella praecipua, Grave, 2004
|
publication ID |
https://doi.org/ 10.5281/zenodo.4689555 |
|
DOI |
https://doi.org/10.5281/zenodo.6267707 |
|
persistent identifier |
https://treatment.plazi.org/id/AF198160-FFBE-FFCC-FF29-99E7FB8C6F11 |
|
treatment provided by |
Felipe |
|
scientific name |
Batella praecipua |
| status |
sp. nov. |
Batella praecipua View in CoL n. sp.
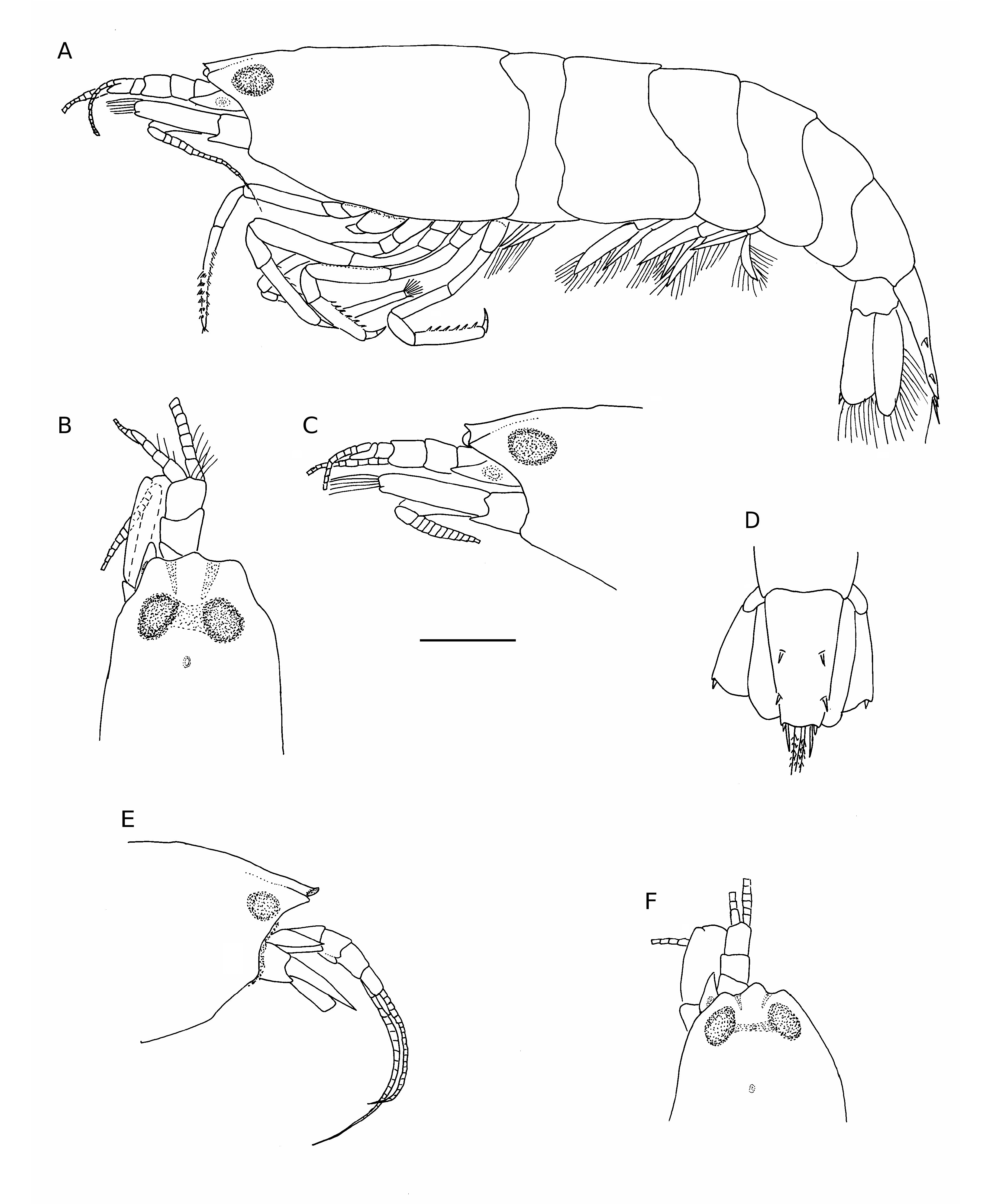
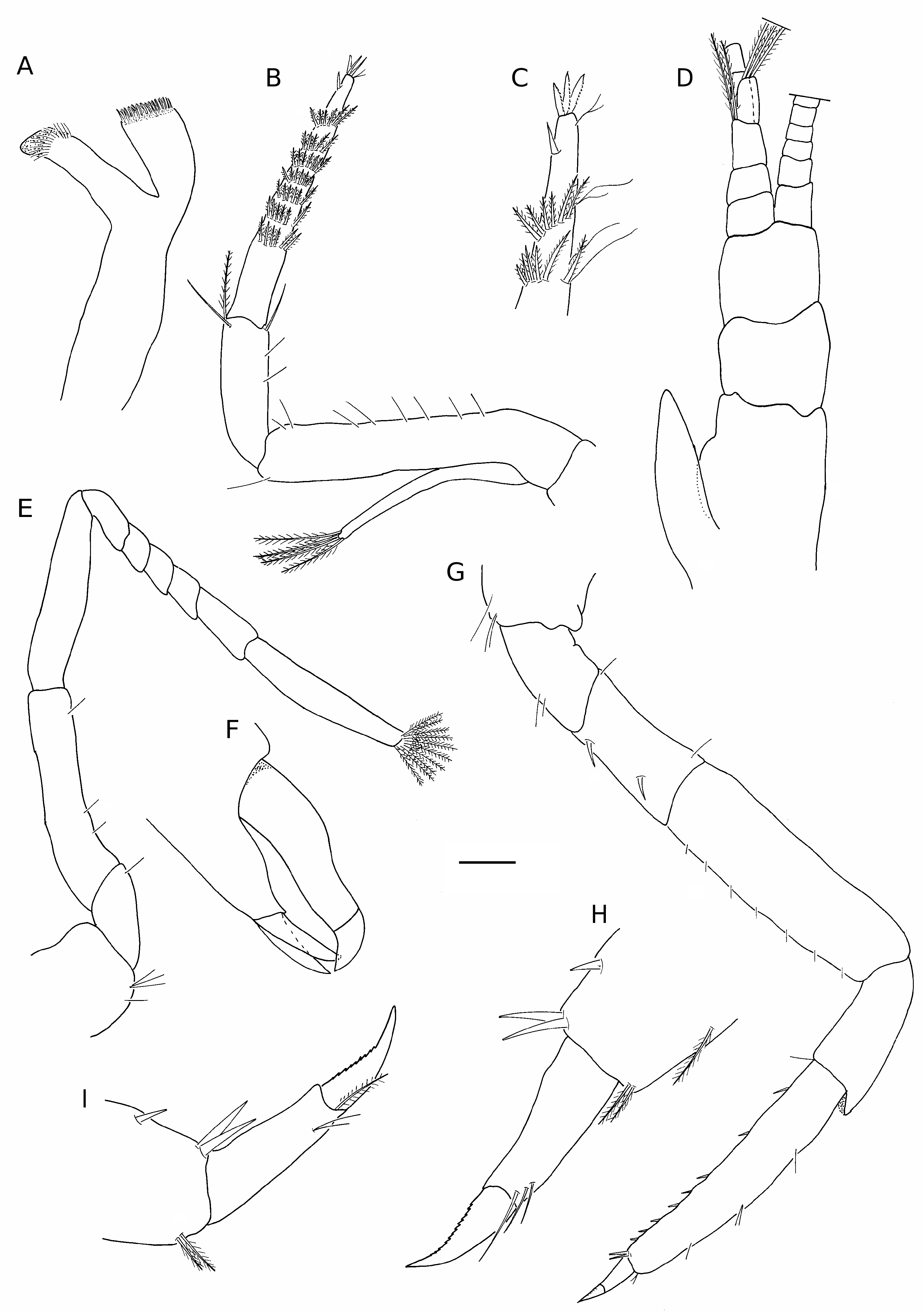
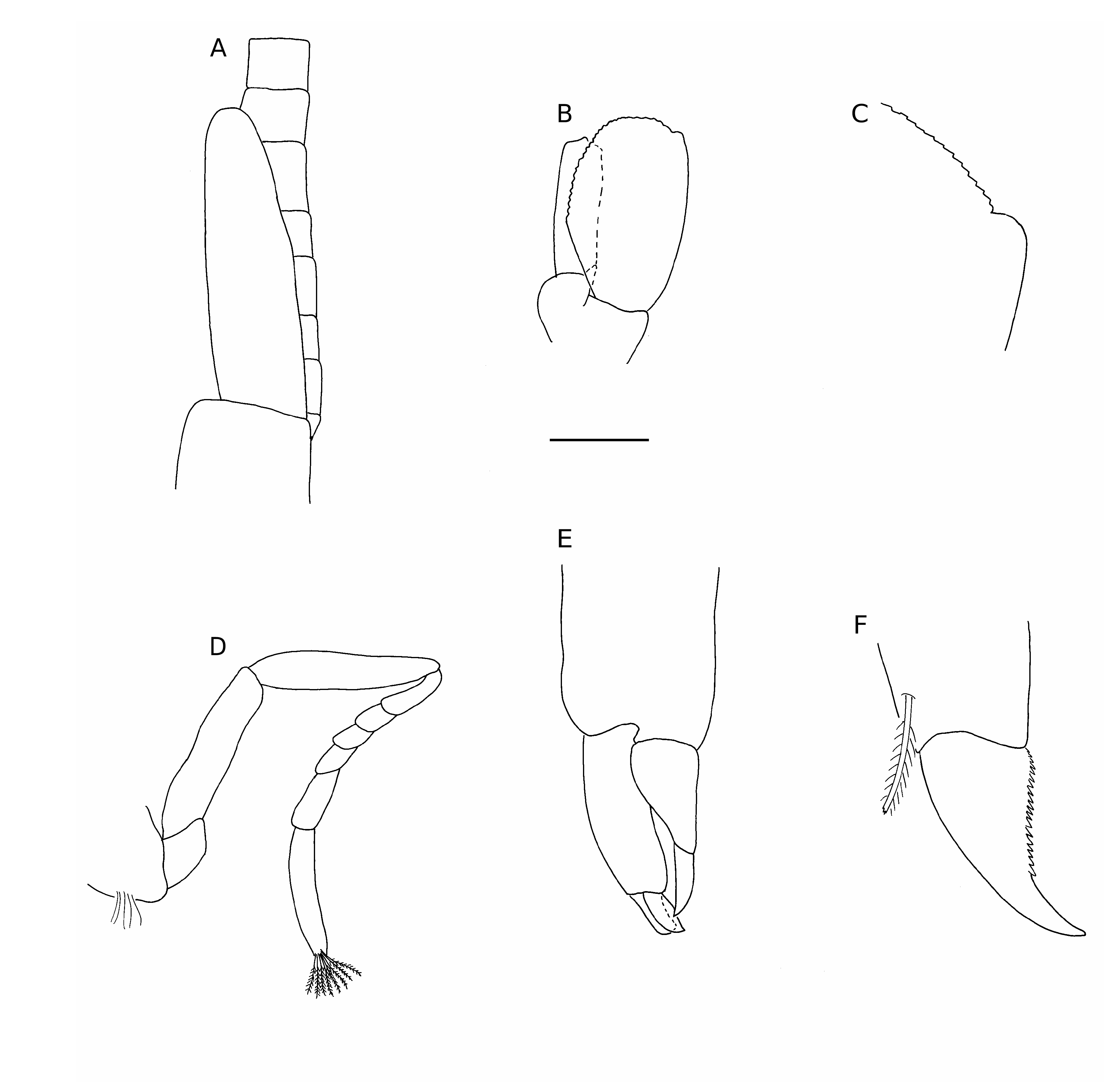
( Figs 1-3 View FIG View FIG View FIG )
TYPE MATERIAL. — Holotype: MUSORSTOM 6, New Caledonia, 22°53’8”S, 167°13’9”E, 425-440 m, “dans une éponge avec des sténopodides”, non ovigerous, post-orbital carapace length (pocl) 5.50 mm ( MHNN Na 11396). Paratype: New Caledonia, 22°52’42”S, 167°11’90”E, 400 m, rocky bottom with sponges, 12. V.1995, leg. A. A. Myers, non ovigerous, pocl 2.60 mm ( OUMNH 2004-01-0001).
ETYMOLOGY. — From the Latin praecipuus, meaning peculiar. In reference to the frontal region of the carapace, exhibiting a non-tridentate structure.
HABITAT. — Known only from New Caledonia, 400- 440 m deep. The holotype was collected with unidentified stenopodidean shrimp from a sponge. No host association is recorded for the paratype specimen, but this latter sample also contained examples of Spongicola levigata Hayashi & Ogawa, 1987 and Periclimenes forcipulatus Bruce,1991 , both known hexactinellid sponge associates.
DESCRIPTION
Carapace smooth, lacking setae ( Fig. 1 View FIG A-C, E, F). Anterior margin trilobed; rostrum poorly developed, tip upturned, adrostral furrows shallow, falling short of eyes. Pterygostomial angle produced, rounded. Ventral margin of carapace straight, not fringed with setae; posterior margin with shallow cardiac notch. Inconspicuous tubercle present on gastric region. Eyes not visible in dorsal or lateral view, anteriorly exposed.
Antennular peduncle ( Fig. 2D View FIG ) short, second and third segments subequal in length, approximately half length of first segment; stylocerite well developed, slightly overreaching first segment, narrowing distally to a blunt point, ventral carina of first article poorly developed, statocyst well developed. External flagellum biramous, outer branch consisting of single segment ( Figs 2D View FIG ; 3A View FIG ), longer free ramus approximately 0.4 times carapace length. Mesial flagellum slender, filiform, approximately 0.7 times carapace length.
Carpocerite of antenna ( Fig. 1A, B, E, F View FIG ) falling short of antennular peduncle; basicerite with well developed ventral tooth; scaphocerite ( Fig. 3B View FIG ) reaching just past antennular peduncle, oval shaped, anterior margin rounded, lateral tooth rounded ( Fig. 3C View FIG ), poorly developed, not reaching past lamina. Flagellum filiform, approximately equal to or slightly longer than carapace length Mouthparts typical for the genus. Mandible ( Fig. 2A View FIG ) deeply bifurcated, palp absent; molar process cylindrical ending in tuft of spines; incisor process spatulate, distal part terminating in tuft of setae. Third maxilliped ( Fig. 2B, C View FIG ) overreaching antennular peduncle; exopod falling short of antepenultimate segment; ultimate segment bearing seven semicircular rows of plumose setae on mesioventral side, single spine subdistally, tip provided with three serrated spines.
First pereiopods lacking in both holo- and paratype.
Second pereiopods ( Figs 2E View FIG ; 3D View FIG ) short, subequal in length to third maxilliped. Ischium and merus elongate; carpus five-segmented, distal segment longest, segment length ratio (proximal to distal) 1.0:0.5:0.5:0.6:1.2. Dactylus ( Figs 2F View FIG ; 3E View FIG ) short, beak-like; 0.15 in length of propodus; terminating in small spine; fixed finger terminating in two blunt spines; fingers with distal tufts of plumose setae.
Third pereiopod ( Fig. 2G View FIG ) robust; ischium with two spines on inferior margin; merus unarmed, provided with row of short, simple setae on mesial margin, 2.1 times length of ischium; carpus unarmed, equal in length to ischium; propodus armed with eight spines along inferior margin and two spines mediodistally, equal in length to merus; dactylus ( Figs 2H View FIG ; 3F View FIG ) slightly curved, approximately 0.25 times length of propodus, unguis clearly demarcated, serrated on inferior margin, not biunguiculate.
Fourth and fifth pereiopods similar to third, unguis ( Fig. 2I View FIG ) serrated on inferior margin, not biunguiculate; fifth pereiopod more slender.
Pleopods of usual alpheid type.
Uropodal endopod reaching to end of telson, exopod slightly shorter, protopod laterally round- ed, exopod with diaresis complete, ending in semi-acute lateral tooth, lateral spine well developed, overreaching distal margin.
Telson approximately 1.3 length of sixth somite, lateral margins straight, narrowing distally; two pairs of dorsal spines, situated at 0.45 and 0.80 of telson length; posterior margin broadly rounded, laterally with two pairs of spines, mesial pair longest, medially with three to five plumose setae.
REMARKS
Although both specimens lack the first pereiopods (a diagnostic feature in Alpheidae ), they clearly belong to the genus Batella , as demonstrated by the lack of a mandibular palp, the incisor of the mandible terminating in setae rather than serrations, and the tufts of long, plumose setae covering the tips of the fingers of the second pereiopods ( Miya & Miyake 1968; Chace 1988). Until now only two species were known in the genus: Batella parvimanus and Batella leptocarpus . The third described species, Batella bifurcata Miya & Miyake, 1968 , was considered a junior synonym of Batella parvimanus by Miya (1984), as its distinctive characters were based on a misinterpretation of Bate’s figures.
The new species can at once be distinguished from the other species by the non-tridentate nature of the frontal part of the carapace, which is clearly tridentate in both B. parvimanus and B. leptocarpus , although damaged in the unique specimen of the latter species ( Chace 1988). Although it was at first assumed that this nontridentate nature was possibly due to damage to the specimens and potential regeneration, further morphological differences are apparent. The unguis of the ambulatory pereiopods is serrate and non-biunguiculate in B. praecipua n. sp. vs non-serrate and biunguiculate in B. parvimanus and B. leptocarpus ; the pterygostomial angle is rounded and only slightly produced in B. praecipua n. sp. vs triangularly produced in the other species; the uropods are near-equal in length to the telson, as opposed to being distinctly shorter than the telson as in the other species.
Batella praecipua View in CoL n. sp. further differs from B. parvimanus View in CoL in having only a single segmented outer branch of the external flagellum on the antennule, this character being unknown in B. leptocarpus ( Chace 1988) View in CoL .
The holotype was collected from a sponge containing stenopodidean shrimp, whilst the sample containing the paratype also contained two specimens of Spongicola laevigata and a single specimen of Periclimenes forcipulatus View in CoL . As both of these species are known associates of hexactinellid sponges, this lends further credence to the hypothesis by Bruce (1988) that Batella View in CoL species are all probably hexactinellid or other sponge associates. The specimens were collected from depths between 400 and 450 m, representing the deepest record so far for the genus.
| MHNN |
Musee d'Histoire Naturalle |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Batella praecipua
| Grave, Sammy De 2004 |
Batella praecipua
| Grave 2004 |
Periclimenes forcipulatus
| Bruce 1991 |
Batella
| Holthuis 1955 |