
Psammoperca waigiensis
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2004.00111.x |
|
publication LSID |
lsid:zoobank.org:pub:242EC75C-52E2-468A-ABD5-C092D7F3908E |
|
persistent identifier |
https://treatment.plazi.org/id/AE3187EB-2F21-FFDD-2D4B-E17E94B4F8B0 |
|
treatment provided by |
Diego |
|
scientific name |
Psammoperca waigiensis |
| status |
|
I Psammoperca waigiensis View in CoL correlated with the broadening of the otic commissure and the lateral extension of the frontal plate above the posterior part of the orbit. This is observed in † Lates bispinosus ( Fig. 9 View Figure 9 ), but the bones are crushed or covered in this area, so that the probable presence of the pedicle cannot be ascertained. † Eolates gracilis specimens do not show any bone in the area; the pedicle is absent.
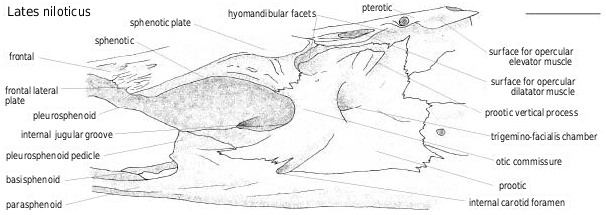
The sphenotic and the prootic share the anterior facet of the articulation of the hyomandibula; the sphenotic and the pterotic share a longitudinal surface for the opercular dilatator muscle, followed by the pterotic oval surface for the opercular levator muscle ( Fig. 7 View Figure 7 ). This pattern is common to perciform fish. However, in some the facets are fused whereas they are always separate in latid fish. The sphenotic lateral plate develops from the anterior facet to the otic commissure level, at which point it is narrower in Tanganyikan species than in other latids (as is the whole neurocranium). The prootic part of the otic commissure anteriorly borders the trigemino-facialis chamber. The chamber is wider in larger individuals, particularly in species with a pleurosphenoid pedicle. A vertical process of the prootic is present, posterior to the hyomandibular anterior facet. On the sphenotic, the otic commissure is ornamented with small pores aligned parallel to the sphenotic anterior border, whereas it is smooth on the prootic.
The pleurosphenoid and the basisphenoid ( Figs 7–9 View Figure 7 View Figure 8 View Figure 9 ) do not show clear differences among Recent latid species and they are not preserved in the examined fossils.
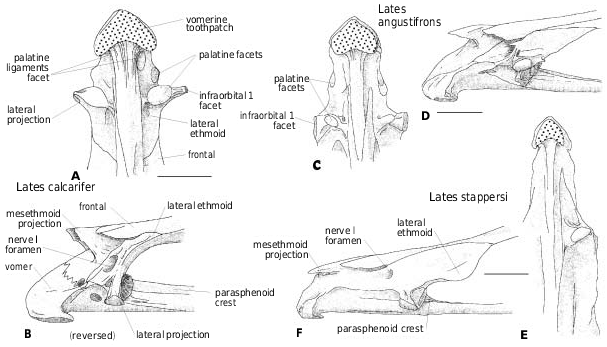
The parasphenoid ( Figs 8 View Figure 8 , 9 View Figure 9 ) ventral border is either straight [ Lates niloticus , L. calcarifer , L. macrophthalmus , † Eolates gracilis , † E. aquensis , † Lates bispinosus ( Figs 8A, B, D View Figure 8 , 9 View Figure 9 )] or slightly curved [ Lates angustifrons , L. longispinis , Psammoperca waigiensis ( Fig. 8C, E, I View Figure 8 )], to strongly bent behind the level of the orbit [ Lates microlepis , L. stappersi , L. mariae ( Fig. 8F–H View Figure 8 )]. The ‘different stages’ of angling recognized by Greenwood (1976) are doubtful and probably a result of the method of drawing (the horizontal level is given by the parasphenoid either in its orbital part or in its braincase part). In transverse section, the parasphenoid ventral face is either flat or slightly concave in the orbital region and convex in the braincase region. A median crest develops on the parasphenoid between the orbits. It starts with a maximal height between the lateral ethmoids and decreases posteriorly ( Figs 2B, D, F View Figure 2 , 8A–H View Figure 8 , 9 View Figure 9 ) and also depending on the size of the individuals. In larger L. niloticus and in L. macrophthalmus , the crest is a low pad. In L. niloticus, Van Neer & Gayet (1988) observed strong variations in the development of the parasphenoid interdigitating suture with the basioccipital and the vomer. The interdigitations are shorter in larger individuals.
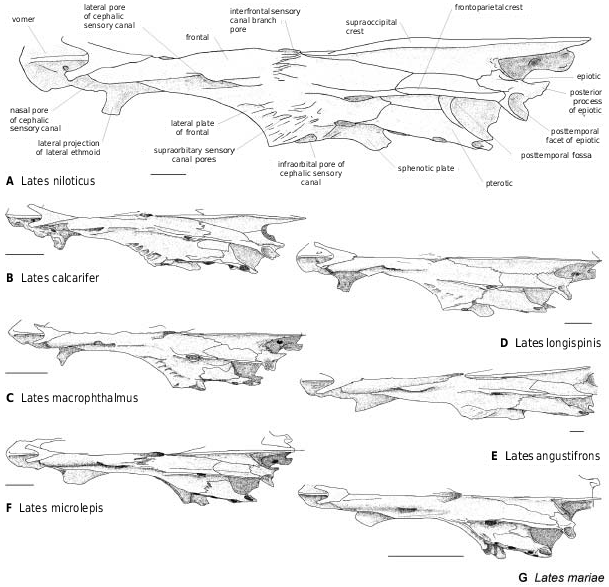
Among fossil latids, the temporal region and the dorsicranium are well known in † Lates bispinosus ( Fig. 9 View Figure 9 ). The posttemporal fossa is opened as in Recent latids, in which, according to Greenwood (1976), the fossa is deep (except Lates stappersi ) and closed off by a tough membrane. The supraoccipital ( Figs 8–10 View Figure 8 View Figure 9 View Figure 10 ) separates the parietals and inserts between the frontals, reaching far anteriorly to the level of the median sensory opening ( Greenwood, 1976). More precisely, it borders from one-quarter to half the frontal length: around one-quarter in Lates calcarifer , L. macrophthalmus and L. mariae ( Figs 8B, D, G View Figure 8 , 10B, D, G View Figure 10 ), one-third in L. longispinis and L. microlepis ( Figs 8C, F View Figure 8 , 10C, F View Figure 10 ), between one-third and a half in L. angustifrons ( Figs 8E View Figure 8 , 10E View Figure 10 ), and a half in L. stappersi and Psammoperca ( Fig. 8H, I View Figure 8 ). These observations, made on few individuals (sometimes a single one), have to be interpreted according to the observations made in L. niloticus . In this species, the larger the individual, the shorter is the extension of the supraoccipital between the frontals (from one-third of the frontal length in small individuals to one-quarter in larger ones). Thus I do not follow Greenwood’s (1976) assumption that in Lates angustifrons , L. mariae and L. microlepis the weakness of the supraoccipital extension is correlated with the ethmoid lengthening. In fact, the development of the pleurosphenoid pedicle may imply that it is the development of the frontal which limits the supraoccipital to the posterior region. Additionally, the elongation of the ethmovomerine region may involve the anterior development of the


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.