
Lampetra soljani, Tutman, Pero, Freyhof, Jörg, Dulčić, Jakov, Glamuzina, Branko & Geiger, Matthias, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4273.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:8CC36A32-5BCA-4BCE-AD6D-DB325C0F5D51 |
|
DOI |
https://doi.org/10.5281/zenodo.6053348 |
|
persistent identifier |
https://treatment.plazi.org/id/AD049877-7415-FFA2-FF1C-2DE5FBA6FE90 |
|
treatment provided by |
Plazi |
|
scientific name |
Lampetra soljani |
| status |
sp. nov. |
Lampetra soljani , new species
( Figs. 3–7 View FIGURE 3 View FIGURE 4 )
Holotype. ZFMK-ICH 103666, 109 mm TL; Bosnia and Herzegovina: side-arm of Neretva River east of Čapljina, 43.106458 17.709536; P. Tutman, 16–17 Jan. 2015.
Paratypes. FSJF 3650, 12 , 105– 126 mm TL; same data as holotype .—FSJF 1037, 1, 111.3 mm TL; Bosnia and Herzegovina: lowermost stream Bregava north of Klepci, 43.052808 17.698089.— FSJF 2184, 1 , 110 mm TL ; Croatia: uppermost stream Norin at Prud, 43.095157 17,619551.
Diagnosis. Lampetra soljani is distinguished from all other species of Lampetra species by having a marmorate flank pattern in live, fully grown ammocoetes, and in some adults (vs. plain colour pattern). It is further distinguished by a combination of characters. Lampetra soljani is most similar to L. lanceolata and L. ninae and is distinguished from these two species by having three velar tentacles (vs. 5 in L. lanceolata , 5-7 in L. ninae ), no posterial teeth (vs. rarely absent, usually a single incomplete row with up to 23 teeth in L. ninae ) and less trunk myomeres (trunk myomeres between last branchial opening and anus 54–57 vs. 57–64 in L. lanceolata , 57–61 in L. ninae ; trunk myomeres between last branchial opening and first dorsal fin origin 28–32 vs. 32–36 in L. lanceolata , 32–35 in L. ninae ; trunk myomeres between last branchial opening and second dorsal-fin origin 42–46 vs. 46–50 in L. lanceolata , 45–49 in L. ninae ).
Lampetra soljani is further distinguished from L. zanandreai by lacking posterial teeth (vs. usually present, very rarely absent). Lampetra zanandreai has a single row of posterials with up to 20 teeth but there are also individuals without posterial teeth ( Hubbs & Potter 1971).
Lampetra soljani is distinguished from L. fluviatilis and L. planeri by having a bicuspid middle endolateral tooth (vs. almost always tricuspid) and the infraoral lamina with 5 teeth (vs. usually 7–9, very rarely 5–6). The Portuguese species L. alavariensis , L. auremensis and L. lusitanica often, but not always, have a bicuspid middle endolateral tooth. Lampetra soljani is distinguished from L. alavariensis , L. auremensis and L. lusitanica by having 54–57 trunk myomeres between the last branchial opening and the anus (vs. 57–63). Lampetra fluviatilis has 58–69 trunk myomeres (n= 883 from more than 14 sites; Hardisty 1986a) and L. planeri has 54–69 trunk myomeres (n=2385 from 18 sites; Hardisty 1986b). However, 54 and 56 are minimum values of ammocoetes just from one site each and usually, the number of trunk myomeres ranges from 57–67 (mean 60–62)
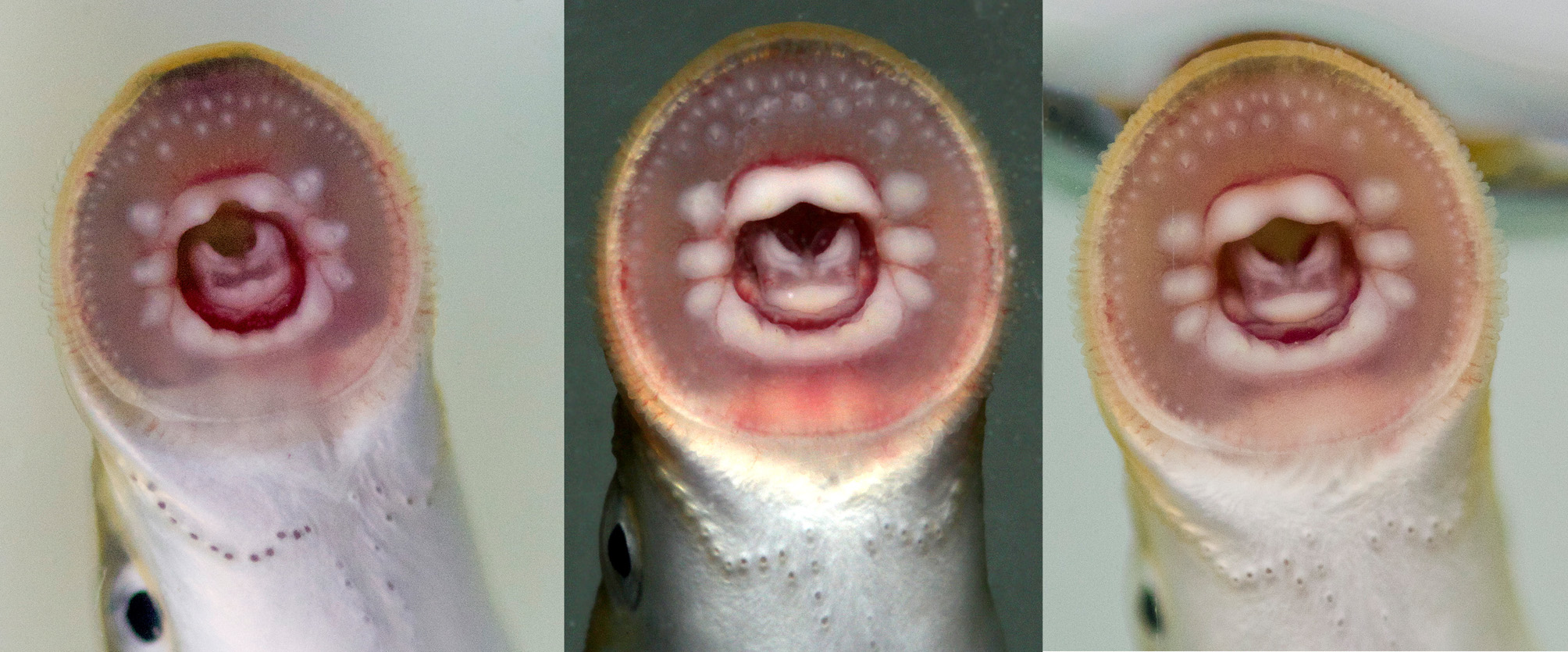
Description. For general appearance see Figs. 3–7 View FIGURE 3 View FIGURE 4 . Morphometric data of pre-spawning adults are provided in Table 2. Oral discs are shown in Fig. 8 View FIGURE 8 . Trunk myomeres between last branchial opening and anus 54–57 (mode 55), between last branchial opening and origin of first dorsal-fin 28–32 (mode 28) and between last branchial opening and origin of second dorsal fin 42–46 (mode 43). Caudal fin spade like. Transverse lingual lamina not fully developed in individuals examined, with 0–4 (mode 2) unicuspid teeth on each side and one enlarged central tooth. In 14 individuals examined for this character, four without teeth on transverse lingual lamina and usually, there are 2–3 lateral teeth, 4 on one side in one individual. Longitudinal lingual laminae straight, each with 4–6 unicuspid teeth, no teeth seen in 12 out of 14 individuals examined. Supraoral lamina with two unicuspid teeth separated by a toothless bridge. Infraoral lamina with 5 teeth, outer two teeth bicuspid, inner three teeth unicuspid. Three bicuspid endolaterals on both sides of oral disc. Anterials in 1–2 rows, usually 1 row, inner row with 4–7 (mode 5 teeth) unicuspid teeth. Exolateral and posterial teeth absent. Three velar tentacles (n=6), middle one shorter than adjacent lateral tentacles, with tubercles on dorsal surface, without wings.
Coloration. Ammocoetes ( Fig. 7 View FIGURE 7 ) are brownish along back with lighter undersides and a marmorate pattern on flank. Tail dark brown. Fins hyaline or pale yellowish-brown. Area around gill openings as well as in front of them reddish from blood vessels. Adults with brown back and upper flank, marmorate in some individuals, belly and lower flank silvery, yellowish in preserved individuals. All fins yellowish. A dark grey blotch near apex of second dorsal fin in some individuals ( Figs. 3–5 View FIGURE 3 View FIGURE 4 ). Lateral-line neuromasts unpigmented on ventral surface and in prebranchial region.
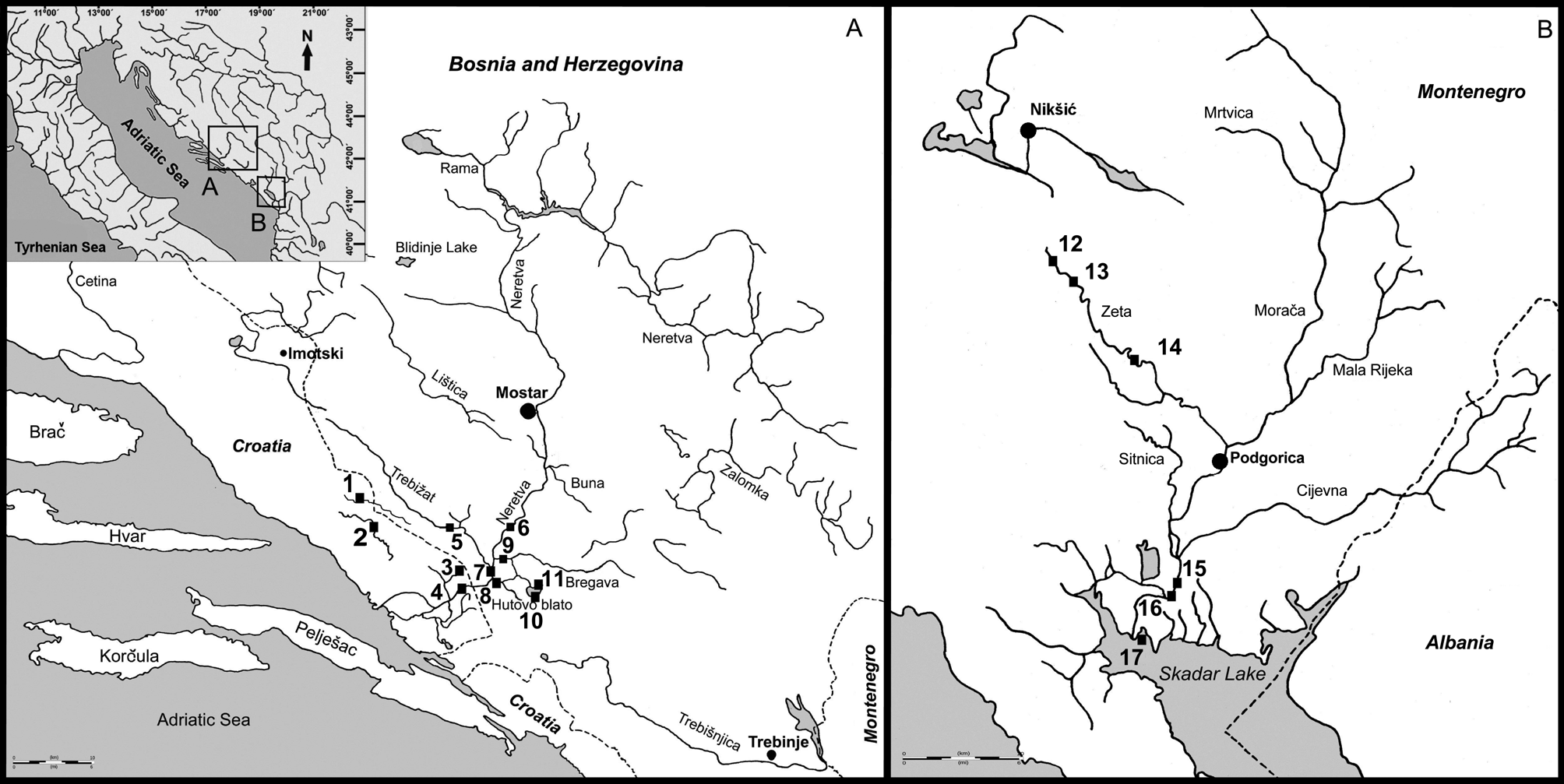
Distribution. Lampetra soljani is only known from the Neretva drainage in Croatia and Bosnia and Herzegovina. Lamprey from the Morača River drainage including Zeta River are preliminary identified as L. soljani based on molecular characters. The Morača River flows to Lake Skadar in Montenegro. The actually known distribution area is shown in Fig. 9 View FIGURE 9 based on records listed in Table 3.
Habitat and biology. Ammocoetes are found in silt, muddy or fine sandy-muddy sediment in river sections with slow or no current, in shallows at banks or backwaters at water depth of about 10–50 cm. Adults and ammocoetes are commonly caught together. First post-metamorphosis individual was found in September (FSJF 2184).
Etymology. Named in honour of Tonko Šoljan (1907–1980) in appreciation of his contribution to the knowledge and development of ichthyology in Croatia and Bosnia and Herzegovina.
Remarks. Lampreys from the Morača River drainage are identified as L. soljani based on the presented molecular characters. No adult individuals were available to us to test, if these agree with the diagnostic characters given here. Therefore, the identification is preliminary and should be tested in the future, when well-preserved adults become available.
Posterial teeth are often deeply embedded in brook lampreys and we were unable to x-ray the mental disc in L. soljani . Due to the availability of so few adult individuals, we also did not apply any clearing and staining methods. Therefore, we cannot full exclude, that deeply embedded posterials, not visible externally, might be present.
Lampreys are still a challenge for taxonomists and a good example for cryptic species exhibiting only few characters that can be reliably measured or counted (no fin rays, no scales, no ossified structures). Furthermore, lampreys show strong allometric growth after metamorphosis ( Krappe 2004; see also differences in size and shape of body and fins in Fig. 5 +6) limiting morphometric studies to individuals in exactly the same phase of life. Krappe (2004) report growth of the disk and shrinkage of the body and tail between 5-15% after metamorphosis for different body proportions in L. planeri . Therefore, the traditional taxonomy in lampreys is mainly based on their dentition and the number of myomeres. Given that brook lampreys do not feed as adults, and hypothesized that they would use their teeth similarly to their predatory congeners, it can be speculated that the variation in tooth formulae and dentition might be a result of relaxed selection pressure. However, this hypothesis deserves a thorough testing and is far beyond the aims of our study. All brook lampreys have less and smaller teeth compared to their predatory congeners ( Renaud 2011; Kottelat & Freyhof 2007). Potentially, all non-feeding species of Lampetra are satellite species of L. fluviatilis and have been isolated at different time periods from this predatory lamprey. It is likely, that all brook lampreys reduce the teeth and the degree of reduction might be related to the time, brook lamprey species or populations are isolated from their predatory congeners.
Nr. Country Drainage Locality Coordinates Source With regard to the paucity of morphological characters compared to bony fishes, the inclusion of molecular genetic characters into standard taxonomic works in lampreys is of great value. A comprehensive, genome wide comparison of a predatory/non-feeding species pair from the same location ( L. fluviatilis with L. planeri ) revealed strong genetic differentiation, despite the existence of shared mitochondrial haplotypes in these two particular populations ( Mateus et al. 2013b). In line with these findings, Rougemont et al. recently (2017) used RAD sequencing for a genome wide screening of SNP in nine different population species pairs in France and demonstrated again a strong genomic differentiation between the two species, but also showed levels of ongoing gene flow to correlate with geographic connectivity. All known brook lampreys included herein show differences in their mitochondrial DNA sequences between them (COI - this study; control region – Pereira et al. 2010; cytB – Mateus et al. 2013a), but no study exists that included the potential predatory sister taxon of the Eastern European species, so we do not know if the genome wide strong differentiation documented in Mateus et al. (2013b) and Rougemont et al. (2017) is the rule or an exception. Interestingly, while our samples of L. soljani show only a single, potentially diagnostic nucleotide substitution (position 460 C vs T) in their DNA barcode region, the four individuals analysed from the Morača River drainage exhibit two additional diagnostic positions, which are unique among all included specimens and can be perceived in the haplotype network ( Fig. 2 View FIGURE 2 ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |