
Psilopini Cresson
|
publication ID |
https://doi.org/10.3853/j.2201-4349.75.2023.1823 |
|
publication LSID |
lsid:zoobank.org:pub:8B1BBD1D-6F2C-4D33-89A0-319C33F71094 |
|
persistent identifier |
https://treatment.plazi.org/id/AC5287BB-FFFB-FF9B-FEB0-F92E966E6F61 |
|
treatment provided by |
Felipe |
|
scientific name |
Psilopini Cresson |
| status |
|
Tribe Psilopini Cresson
Psilopini Cresson, 1925: 241 (as Psilopinae). Type genus: Psilopa Fallén, 1823 .—Mathis & Zatwarnicki, 1995: 30–50 [world catalog].
Heringinae Enderlein, 1934: 191. Type genus: Heringium Enderlein, 1934 (= Clanoneurum Becker, 1903 ).
Clanoneurinae Enderlein, 1936: 168. Type genus: Clanoneurum Becker, 1903 .
Risini Papp, 1977: 188 (as Risidae ). Type genus: Risa Becker, 1907 View in CoL syn. nov.
Diagnosis. The tribe Psilopini is distinguished from Discomyzini and other tribes of Ephydridae by the following combination of characters: Small to moderately small shore flies, body length 1.50–2.50 mm; usually mostly black, shiny, microtomentum usually sparse, especially on frons and scutum.
Head: Ocellar seta aligned behind anterior ocellus, sometimes only slightly so; reclinate fronto-orbital seta inserted behind, sometimes weakly developed or lacking, lateroclinate (sometimes well developed, divergent, and slightly proclinate), usually much less than ½ length of ocellar seta; arista with 7–14 dorsal rays; conformation of face variable, usually mostly smooth, sometimes distinctly carinate, shallowly pitted or rugose; gena, including midportion, setulose, its posterior margin rounded, mouthparts sometimes moderately to distinctly geniculate, length of labellum equal to haustellum.
Thorax: Mesonotum generally sparsely microtomentose to bare, subshiny or shiny, although variable; supra-alar seta usually lacking, if present, well developed, subequal to postalar seta; arrangement of acrostichal setulae variable; prescutellar acrostichal setae usually present, widely set apart and aligned anterior of posteriormost dorsocentral setae; scutellum usually sparsely setulose; anterior and posterior notopleural setae inserted at about the same level. Wing hyaline to milky white; costal length variable; crossvein dm–cu sometimes lacking.
Abdomen: Male terminalia: Epandrium and cerci distinctive; gonostylus well developed, rarely reduced in some Psilopa Fallén ; subepandrial plate present, this plate fused in some genera with pregonite; pregonite elongate, setulose; postgonite small, bearing 1–6 setae (usually 3); aedeagus ovate in ventral view; ejaculatory apodeme lacking; phallapodeme narrow, triangular in lateral view; hypandrium broad, medially incised.
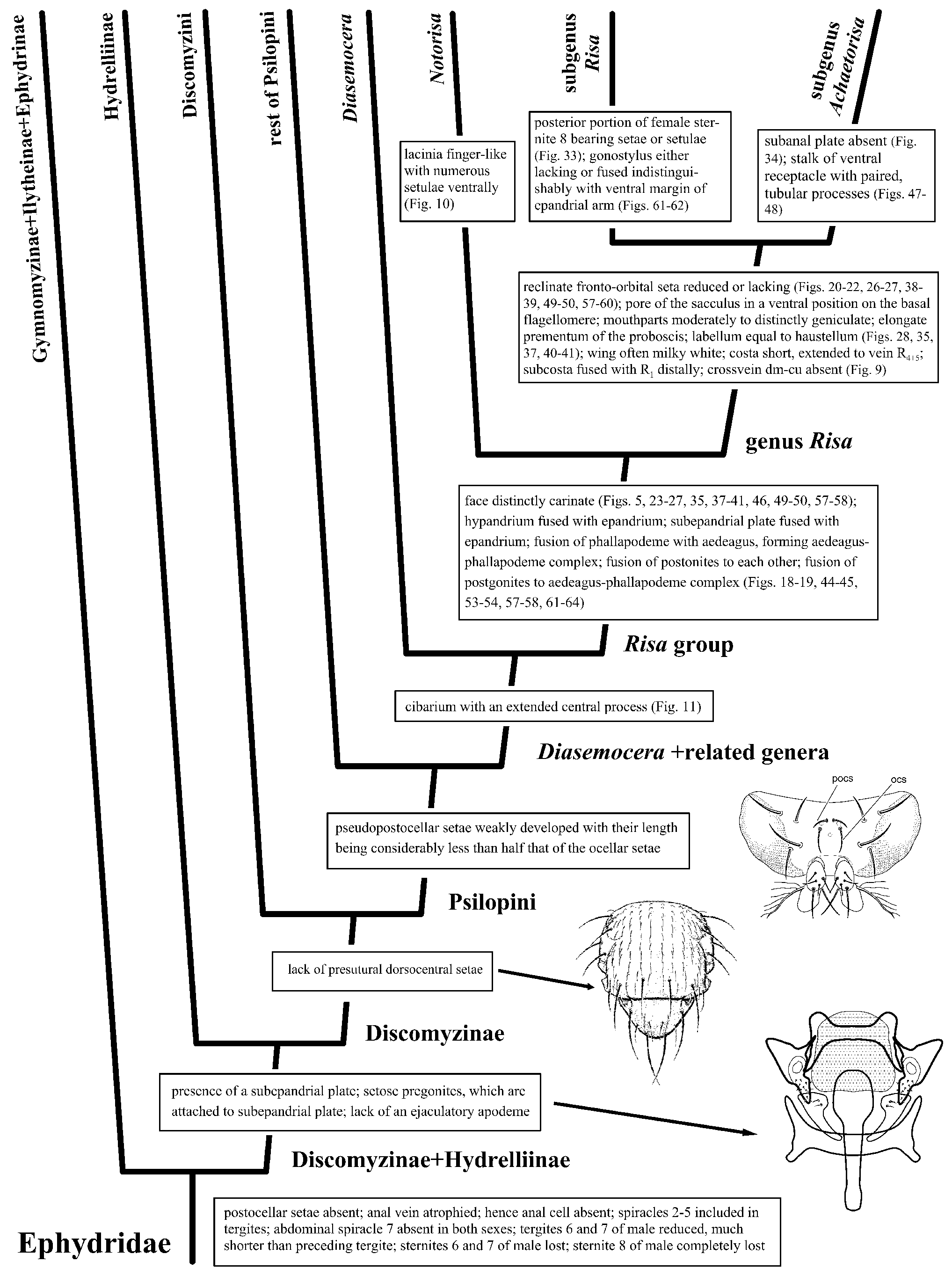
Discussion. The position of the Risa group of genera within the Ephydridae is supported by the following characters: Head: Postocellar setae absent. Thorax: Anal vein atrophied; hence anal cell absent.Abdomen: Spiracles 2–5 included in tergites; abdominal spiracle 7 absent in both sexes; tergites 6 and 7 of male reduced, much shorter than preceding tergite; sternites 6 and 7 of male lost; sternite 8 of male completely lost ( Fig. 1 View Figure 1 ).
Structures of the male terminalia of the Risa group include a few characters that are also found apparently in the shore-fly tribe Atissini ( Ephydridae : Hydrelliinae ). Externally, moreover, there is some resemblance also with specimens of Gymnomyzini ( Ephydridae : Gymnomyzinae ), and the primitive type of cibarium and lack of presutural dorsocentral setae suggest placement in Discomyzinae or Gymnomyzinae . However, the presence of a subepandrial plate, setose pregonites, which are attached to subepandrial plate, and lack of an ejaculatory apodeme are more typical for Discomyzinae , and like other genera of Psilopini , the pseudopostocellar setae are weakly developed with their length being considerably less than half that of the ocellar setae and the stem of the radial vein lacks setulae.
The tribe Psilopini is closely related to Discomyzini, and these two tribes constitute the subfamily Discomyzinae ( Fig. 1 View Figure 1 ). Worldwide, the tribe Psilopini comprises 14 genera and 159 species (Mathis & Zatwarnicki, 1995 and electronic updates). As noted in the above synonymy, we place the tribe Risini as a junior synonym of Psilopini and refer to the taxa formerly placed in Risini as the Risa group of genera. Like other genera of Psilopini , the pseudopostocellar setae are weakly developed with their length being considerably less than half that of the ocellar setae and the stem of the radial vein lacks setulae.
In this study, we include both external and internal morphological characters, and based on this assessment, we suggest that the Risa group of genera is a monophyletic lineage that is also supported by several hypopygial characters: hypandrium fused with epandrium; subepandrial plate fused with epandrium; fusion of phallapodeme with aedeagus, forming aedeagus-phallapodeme complex; fusion of postgonites to each other; and fusion of postgonites to aedeagus-phallapodeme complex.
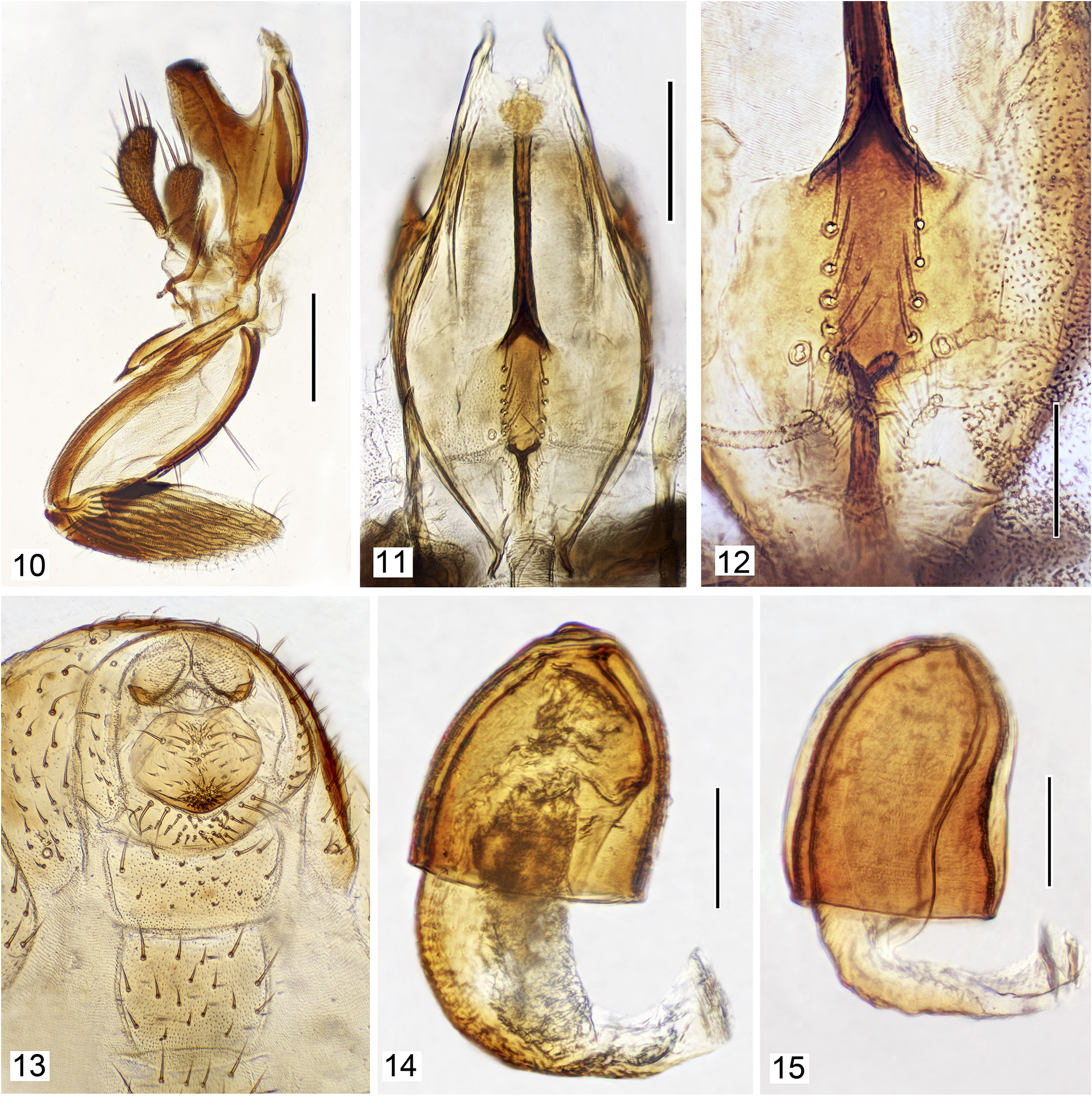
Within the tribe Psilopini , the Risa group is closely related to the genus Diasemocera Bezzi. This conclusion is based on the following characters: Seven pseudotracheae ( Fig. 10 View Figures 10–15 ) and cibarium that are similarly shaped and with an extended central process ( Figs 11, 12 View Figures 10–15 ). Other genera within Psilopini , such as Clanoneurum Becker , Cnestrum Becker , and Trimerina Macquart , have seven pseudotracheae, but these genera have a simple cibarium. Only Diasemocera has an elongated cibarial process ( Zatwarnicki, 2018).
The other genera referred to above ( Clanoneurum , Cnestrum , Diasemocera , and Trimerina ) also have an association with the plant family Amaranthaceae . For example, D. leucostoma (Meigen) has been reared from Beta vulgaris L., and most species of Diasemocera breed in saline habitats where plants of the genera Chenopodium L., Seidlitzia L., and Suaeda Forsskål proliferate. We acknowledge, however, that specific host-plant information is not presently available for many species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Psilopini Cresson
| Mathis, Wayne N. & Zatwarnicki, Tadeusz 2023 |
Risini
| Papp, L. 1977: 188 |