
Schistura papulifera, Kottelat, Maurice, Harries, Daniel R. & Proudlove, Graham S., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.175283 |
|
DOI |
https://doi.org/10.5281/zenodo.5663813 |
|
persistent identifier |
https://treatment.plazi.org/id/AB5D87EC-2A62-BF78-FF07-FD6BFD3951DA |
|
treatment provided by |
Plazi |
|
scientific name |
Schistura papulifera |
| status |
sp. nov. |
Schistura papulifera View in CoL , new species
( Fig. 1 View FIGURE 1 )
Holotype. MHNG 2680.074 (45.1 mm SL); India: Meghalaya, Maintia Hills, Krem Umsngat entrance (25º11’14”N, 92°21’03”E) to Synrang Pamiang cave system (main entrance 25º12’48”N, 92°21’48”E); Meghalayan Adventurers Association, Shillong, Feb. 1999.
Paratype. CMK 16040, 1 (48.7 mm SL); same data as holotype.
Diagnosis. Schistura papulifera is distinguished from all other species of the genus in having the lower half of the head covered by numerous small skin projections and 5 pores in the supratemporal canal of the cephalic lateralline system. In addition, the body is white, the eyes are vestigial, the lateral line is incomplete (reaching only to vertical above pelvicfin origin) and there is no axillary pelvic bone.
Description. See Figure 1 View FIGURE 1 for general appearance and Table 1 View TABLE 1 for morphometric data of holotype and paratype.
A moderately elongate nemacheiline loach, with body depth slowly increasing up to a point slightly in front of dorsalfin origin, then decreasing to caudalfin origin. Head depressed, body slightly compressed anteriorly to compressed posteriorly. Pectoral fin not reaching pelvicfin base. No axillary pectoral lobe. No axillary pelvic lobe. Pelvic fin not reaching beyond anus; origin under base of first to second branched dorsalfin rays. Distance between anus and analfin origin about equal to onethird of snout length. Caudal fin deeply emarginate. Ventral and dorsal adipose crests on caudal peduncle, which is 1.4 times longer than deep. Distal margin of dorsal fin convex.
D. 4/8½; A. 3/56½; C /9+8/; P. 1/1112; V. 1 /7.
Body entirely covered by scales, except on belly between pectoralfin bases. Lateral line incomplete, with about 12 pores, reaching posteriorly to vertical over pelvicfin origin. Cephalic lateralline system with 6 supraorbital, 3+12 infraorbital, 9 (? very difficult to count) preoperculomandibular, and 5 supratemporal pores.
Anterior nostril pierced on anterior side of a long, pointed flaplike tube not reaching eye. Mouth gape about 1.5 times wider than long ( Fig. 2 View FIGURE 2 a). Lips fleshy, covered by small folds and numerous transverse rows of small papillae (possibly unculi). No median incision in upper lip. A median incision in lower lip. Processus dentiformis present. A shallow median concavity in lower jaw. Inner rostral barbel reaching slightly beyond corner of mouth; outer rostral and maxillary barbels reaching beyond half of postorbital length of head. Intestine with a small loop immediately behind (and adjacent to) stomach ( Fig. 2 View FIGURE 2 b). Air bladder without free posterior chamber (in dissected paratype).
In both specimens eye vestigial, subcutaneous, and externally appearing as a small diffuse blackish spot, not communicating with outside by a canal. Lower half of head covered by numerous soft skin projections, appearing as bumps or blunt tubercles ( Fig. 3 View FIGURE 3 ). No apparent sexual dimorphism.
Coloration. Preserved specimens white, with a few scattered flecks of black pigment on dorsal fourth of body. In life, white and appearing eyeless.
Etymology. From the Latin papula (a small rounded tumor on skin) and ferre (to bear on oneself). An adjective.
Remarks. The shape of the caudal fin appears different in the two specimens, but in the larger one close examination reveals that it has been bitten or mutilated and regenerated, a common observation in cavernicolous benthic fishes.
Relationships. The Synrang Peninsula cave system is located on the southern slope of the Jaintia Hills and drains southwards to the Meghna River in Bangladesh. The Meghna River is a tributary of the Brahmaputra River, which it joins in the Ganges delta, which constitutes most of the lowlands of Bangladesh.
Although the Ganges delta and adjacent areas are some sort of birthplace for Asian ichthyology, with the founding work of Hamilton (1822), our state of knowledge of its fish fauna is extremely frustrating. Despite the steady flow of publications, monographs and other scholarly and popular treatises, we are desperately lacking sound reviews based on largescale fieldwork and decently preserved material. This explains why discussion of the relationships of S. papulifera with surface species can hardly be made.
Menon (1987) reviewed the Indian nemacheilines and recorded Nemacheilus sikmaiensis Hora , N. barapaniensis Menon and N. reticulofasciatus Singh and Banarescu (in Singh et al., 1982) from eastern Meghalaya (despite the title of his book, Menon did not examine material from Bangladesh). Schistura papulifera is distinguished from them and from all species of nemacheiline loaches by having the lower half of the head covered by small skin projections (vs. no such projections) and 5 pores in the supratemporal canal of the cephalic lateralline system (vs. 3, except in Yunnanilus , which has 4). Furthermore, S. papulifera is distinguished from the epigean species by absence of color pattern and possession of vestigial eyes.
Nemacheilus sikmaiensis View in CoL of Menon (1987) was placed in the genus Schistura View in CoL by Kottelat (1990: 208), who commented that records of the species from outside the Irrawaddy drainage (that is, the Brahmaputra drainage) require confirmation. Although the name appears regularly in the Indian literature, until recently no attempt had been made to critically compare material from the two drainages. Vishwanath and Laisram (2001) compared populations from the Brahmaputra and Irrawaddy drainages in Manipur, and concluded that they are conspecific. Besides superficial differences such as the absence of pigmentation pattern and general appearance, S. papulifera View in CoL is distinguished from S. sikmaiensis View in CoL in having a distinct processus dentiformis (vs. absence), no median incision in upper lip (vs. presence), absence of an axillary pelvic lobe (vs. presence), and an incomplete lateral line reaching about on a vertical through pelvicfin origin (vs. complete).
Nemacheilus barapaniensis Menon 1987 View in CoL is a replacement name for Physoschistura elongata Sen and Nalbant View in CoL (in Singh et al., 1982) because it is a junior homonym of Aborichthys elongatus Hora View in CoL , if both are placed in Nemacheilus View in CoL . As discussed by Kottelat (1990), Aborichthys View in CoL and Physoschistura View in CoL are distinct and valid genera. Even if there are uncertainties about the actual generic position of P. elongatus View in CoL , in any case it is not a Nemacheilus View in CoL as defined by Kottelat (1984, 1990). Following the International Code of Zoological Nomenclature (art. 59.4), a junior homonym rejected after 1960 (here Physoschistura elongata View in CoL rejected by Menon, 1987) must be reinstated as valid if it is later considered not congeneric. Nemacheilus barapaniensis View in CoL therefore is an objective junior synonym of Physoschistura elongata View in CoL , without nomenclatural validity.
In the original description of P. elongata, Banarescu and Nalbant View in CoL (in Singh et al., 1982) diagnosed Physoschistura View in CoL mainly by the presence of a free posterior chamber of the air bladder. Kottelat (1990: 77) commented that this character alone does not define a lineage as it appears in other nemacheiline species and groups that do not appear closely related to Physoschistura View in CoL . On the other hand, several species share the short, somewhat massive appearance of the type species, P. brunneanus Annandale View in CoL , and a particular lip construction which Kottelat (1990: 77) used to rediagnose the genus. Due to the limited number of specimens available (usually 1 or 2 for each species), it was not possible to dissect individuals of these species to check whether the airbladder was present. Furthermore, Banarescu and Nalbant do not indicate the number of specimens of each species they checked for this character and what variation they observed. This reduces the usefulness of this character.
The relatively arched mouth of S. papulifera and its fleshy lip, especially the fleshy median pads of the lower lip (also with numerous small folds), is similar to that of species of Physoschistura , and we considered the possibility that it might belong to that genus. The air bladder does not have a free posterior chamber in the single dissected specimen, and the general appearance of S. papulifera (relatively slender body, with slightly depressed head) differs from that of species placed in Physoschistura by Kottelat (1990) (relatively deep and compressed head and body; see photographs in Kottelat, 1990), and we prefer to retain it in Schistura . Schistura is conveniently a catchall genus within which some lineages can be recognized, but we are still far from a broad general knowledge of its taxonomic diversity. The “genus” is very speciose in hilly areas of southeastern Asia (more than 100 species have been described in the last 15 years; Kottelat, 1990, 1998, 2001; Freyhof and Serov 2001; Zhu 1989), and many more should be expected. In addition, many of the species from the northwestern part of the Indian subcontinent are probably polyspecific assemblages. When this area is better surveyed, when better preserved specimens become available, when the species are more critically described, and when the material becomes accessible to the international scientific communities, it will probably be possible to recognize affinities among surface species (across political boundaries), and it could be possible to identify surface species related to S. papulifera . Today we do not even know which (if any) species occur in the same drainage in Meghalaya.
Nemacheilus reticulofasciatus View in CoL of Menon (1987) was originally described as Mesonoemacheilus reticulofasciatus View in CoL . The genus Mesonoemacheilus View in CoL includes species restricted to peninsular India (Kottelat, pers. obs.), and M. reticulofasciatus View in CoL belongs to a generally southeast Asian lineage (presently included in Schistura View in CoL s.l.) diagnosed by the shape and position of the black spot at the caudalfin base, the body pigmentation pattern consisting of very thin bars anteriorly, more or less connected at their upper extremities and broad bars posteriorly. This lineage includes, among others, S. vinciguerrae View in CoL , S. poculi View in CoL , S. sikmaiensis View in CoL (Kottelat, in prep.). Schistura papulifera View in CoL is distinguished from S. reticulofasciata View in CoL (as figured by Singh et al., 1982) by the general body shape, the latter having a deeper body, shorter head, and a much stouter caudal peduncle (we have not attempted to quantify these differences as we do not know which body landmarks Singh et al. used for morphometric analyses; the caudal peduncle appears clearly longer than high in their figure, whereas the values in the text suggest the reverse).
A synopsis of the fish fauna of Meghalaya ( Sen, 1995) conveys little useful information, except that the following species of Schistura View in CoL are reported from the Jaintia Hills: S. multifasciata (Day) View in CoL , S. reticulofasciata View in CoL , S. savona (Hamilton) View in CoL , and S. sikmaiensis View in CoL . Rahman (1989) recorded the following species of Schistura View in CoL from Sylhet District of Bangladesh (adjacent to eastern Meghalaya and downstream in the Meghna basin): S. zonata M'Clelland View in CoL , S. beavani (Günther) View in CoL , S. sikmaiensis View in CoL , and S. savona View in CoL . At this stage, and for reasons mentioned above, it serves no purpose to discuss further how each of these species (assuming they are correctly identified) differs from S. papulifera View in CoL . All have a pigmentation pattern and normally developed eyes, and none has the lower half of the head covered by small skin projections and the 5 pores in the supratemporal canal of the cephalic lateral line system, which are diagnostic of S. papulifera View in CoL .
Hora (1924: 28) recorded a nemacheiline population from Siju Cave in the Garo Hills (Meghalaya). He later ( Hora, 1935: 63) identified it as Nemacheilus beavani . Menon (1987: 175) described S. sijuensis on the basis of material from the entrance to the cave, and stated without further comments that a "specimen found 1700 ft. from the entrance is uniformly pale in colour". He examined 2 specimens from inside the cave, but provided no information on the second one. These are apparently the two specimens described and figured by Hora (1924), who reported that they have remains of a pigmentation pattern. Hora's figures show a fish from outside the cave and one from 1700 ft inside with differences in body shape, which suggests that two species might be involved. It has never been discussed and demonstrated that they are conspecific. In fact, the differences in head shape, eye size, etc., which Hora attributed to cave adaptation, might as well be specific characters. It is difficult to comment on the identity of the cave specimens; they seem to be discolored specimens of a surface species, but we have no information on the surface species in the immediate vicinity. Siju Cave (25°21’02"N, 90°41’04"E) is located in the South Garo Hills in the western part of Meghalaya. Siju is over 170 km west of Synrang Pamiang, and the two cave systems lie almost at opposite ends of the band of cavebearing limestone that runs along the southern boundary of the Meghalayan plateau. Given the southern trend of drainage in the area and likely discontinuities in the limestone band, it is unlikely that there can be any hydrological connection between Siju Cave and Synrang Pamiang. It does not seem necessary to elaborate further on the (non) relationships between the two cave species, especially in the absence of material.
Habitat and field observations. Schistura papulifera is currently known with certainty only from the Krem Synrang Pamiang system, in the Jaintia Hills, eastern Meghalaya ( Fig. 4 View FIGURE 4 ). The Jaintia Hills lie in the eastern part of the Indian state of Meghalaya. The Synrang Pamiang cave system is located about 1 km east of the village of Chiehruphi. This village lies on the NH44 road that connects Sonapur, in the south, to Kliehriat (Jowai), in the north, and is approximately 5 km north of the village of Lumshnong. The entrances of the cave are to the north of the track that connects Chiehruphi to the neighbouring village of Musianglamare.
Synrang Pamiang is an extensive system, with over 14 km of mapped cave passages and five known entrances. The main drain of the system is a large meandering streamway that extends for over 7.5 km before issuing to the surface near the Krem Khlieh Trai Lum entrance (25°10’23"N, 92°21’20"E). The water then flows southwards in the basin of the Meghna River, a main tributary of the Brahmaputra in Bangladesh. The cave is in nummulitic Eocene limestone (Lakadong, Lower Sylhet). Exploration and mapping were conducted by the Meghalayan Adventurers Association and various European speleologists in February 1997, 1998, and 1999.
Fish were only observed in a still gour (rimstone) pool with sandy bottom, 1.5 x 7 m, 1 m deep. In February, the passage carries no water, but would do so in the wet season (May–October). Up to 20 individuals were seen on each of five trips to the area. They exhibited no response to light, but were disturbed by movements of the water (A. Boycott, pers. comm.).
Other fauna recorded in Synrang Pamiang included a slug (Pulmonata), a pseudoscorpion (Pseudoscorpiones), spiders ( Heteropoda robusta and Oonops sp.), clumps of harvestmen (Opiliones), troglomorphic harvestmen (Opiliones), shrimp ( Macrobrachium sp.), crabs ( Paratelphusa sp.?), woodlice (Oniscoidea, Philoscia sp.?), millipedes ( Trachyiulus mimus ), springtails (Collembola spp, including a troglomorphic species), brown crickets ( Rhaphidophoridae ), troglomorphic cockroaches, beetles ( Cholevidae ), flies ( Mycetophilidae ), a gecko ( Gekkonidae ) and occasional bats (Microchiroptera) ( Gebauer, 2003; C. Fischer, pers. comm.). In general terms this is typical of many other caves in the Jaintia Hills. Spiders of the genus Heteropoda are routinely recorded from the entrance areas of caves, and appear to be present throughout Meghalaya. Darkly pigmented opilionids are often seen in large aggregations on overhanging rocks close to cave entrances, and troglomorphic species are occasionally recorded farther into the caves. Large shrimp ( Macrobrachium sp.) appear to be ubiquitous in the pools and streamways of the Jaintia caves. Crabs ( Paratelphusa sp.?) are common in many caves and build burrows in moist gravel on cave floors. Woodlice are frequently encountered, especially near decaying vegetation deposited by monsoon floods. The millipede Trachyiulus mimus has been noted in most of the caves visited in the Jaintia Hills, and was originally described from Siju Cave in the Garo Hills. Large brown rhaphidophorid crickets are found in most caves. These are yet to be formally identified, but may turn out to be a species of Diestrammena described from caves in the Jaintia by Chopard (1924). A troglomorphic cockroach, Spelaeoblatta caeca , has been described from the Jaintia caves ( Chopard, 1921). Bats are frequently noted in the caves and Hipposideros appears to be a common genus. It seems that the gecko and slug were accidental trogloxenes recorded near an entrance.
The stomach of the dissected paratype contained no identifiable remains.
Simon Brooks (pers. comm.) reports that fishes very similar in appearance to S. papulifera were seen in another cave, Krem Wah Ryngo, in the Sutnga area 30 km NE of Krem Synrang Pamiang. Their habitat was similar. They have subsequently been recorded from several other caves in the Sutnga area, and these observations are summarized in Table 2.
These records refer to observations of S. papulifera or a very similar species of strygobitic nemacheline. The records were made within a day or two of the observations, and the observers were shown photographs of the fish and asked to confirm that it matched their observations.
Apart from Schistura papulifera , there are two other fish taxa that are commonly recorded in the caves. Glyptothorax sp. (50–80 mm) appear to frequent areas with faster flowing water. A large (20–30cm) cyprinid is found in larger pools and deep streamways. From photographs, we identify it as a species of Tor or Neolissochilus . These are often very pale, but have well developed eyes and are epigean fish.
During the monsoon season (May to October), Meghalaya is subjected to extremely high rainfall. During February the water levels in the caves are relatively low, but it is evident that many of the lower level passages carry substantial watercourses in the wet season. Vegetation and other flood debris can be found deep within the caves, and many passages contain large gravel pools of standing water. Schistura papulifera is typically found in such pools of standing water, rather than in flowing streamways. It appears to be most abundant in areas where the pools are both deep and extensive. It is likely that it retreats to such areas as the onset of the dry season results in falling water levels, and that the larger pools offer the best resources in terms of food and relatively stable oxygen levels.
TABLE 2. Observation of Schistura species (probably S. papulifera ) in caves in the Sutnga area.
Cave Name Coordinates Date Notes
Krem Shynrong 25°21’01.2"N Feb 2001 In most parts of lower series of passages where pools of
Labbit 92°30’10.5"E D. Harries standing water were present. Particularly abundant in large deep canals of standing water. Attracted to meat bait.
Krem Risang 25°19’54.0"N Feb 2001 Common in pools of standing water in lower series of the
92°29’46.0"E D. Harries cave.
Krem Umthloo 25°19’33.1"N Feb 2001, 2002, Widespread in cave system, very abundant especially in
92°30'45.5"E 2003 & 2005 upstream part of cave. D. Harries
Krem Liat Prah 25°22’31.5"N Feb 2003 Uncertain record. 92°32’18.6"E N. Robertson
Krem Krang 1 25°20’21.5"N Feb 2003 Abundant in pools of streamway. 92°30’13.7"E D. Harries
Krem Wah 25°20’38.0"N Feb 2003 Photographed.
Shyngktat 92°30’25.6"E A. Neumann

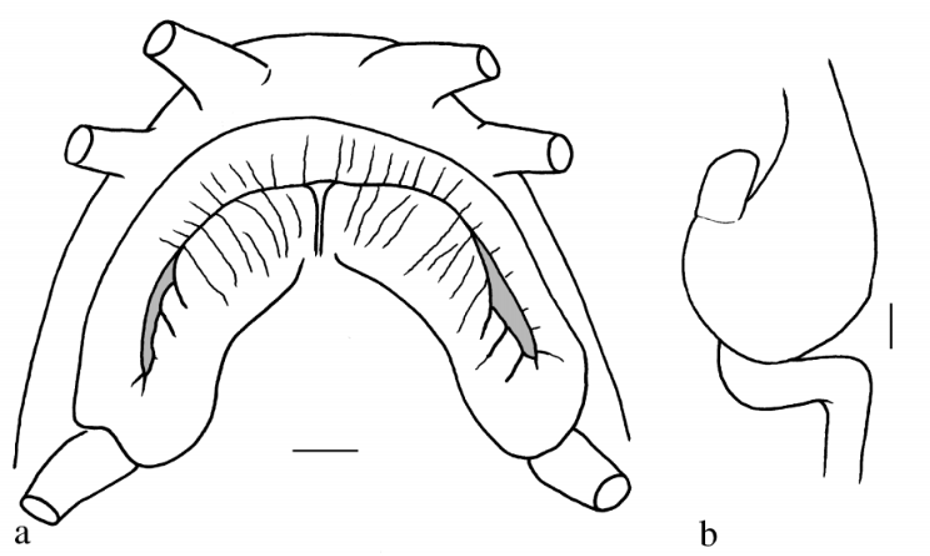
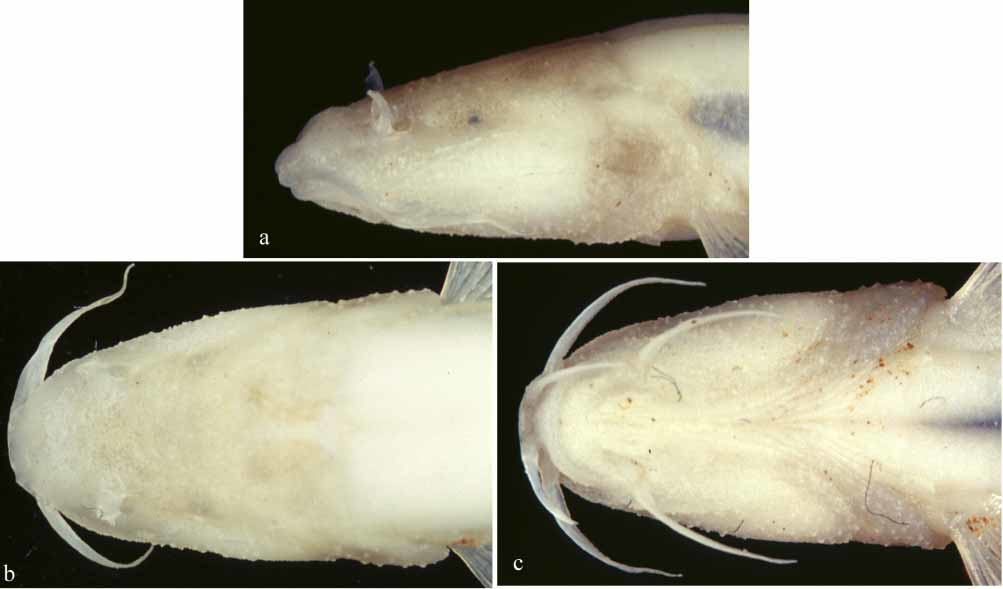

TABLE 1. Morphometric data of holotype (MHNG 2680.074) and paratype (CMK 16040) of Schistura papulifera.
| holotype | paratype | |
|---|---|---|
| % SL % HL | % SL % HL | |
| Standard length (mm) | 45.1 | 48.7 |
| Total length | 122.4 | 121.4 |
| Dorsal head length | 22.4 | 24.8 |
| Lateral head length | 25.9 116 | 27.1 109 |
| Predorsallength | 57.0 | 57.7 |
| Prepelviclength | 55.4 | 56.1 |
| Preanus length | 76.3 | 73.1 |
| Preanal length | 79.8 | 79.1 |
| Head depth | 11.3 50 | 11.5 46 |
| Body depth (D) | 13.1 | 13.3 |
| Depth of caudal peduncle | 10.2 | 10.1 |
| Length of caudal peduncle | 14.6 | 14.4 |
| Snout length | 11.3 50 | 13.1 53 |
| Maximum head width | 14.9 66 | 15.0 60 |
| Body width at dorsal origin | 10.6 | 9.5 |
| Interorbital width | 9.3 42 | 8.6 35 |
| Length of pelvic fin | 17.3 | 15.6 |
| Length of pectoral fin | 21.1 | 18.5 |
| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Schistura papulifera
| Kottelat, Maurice, Harries, Daniel R. & Proudlove, Graham S. 2007 |
Nemacheilus barapaniensis
| Menon 1987 |