
Parazhelestes robustus, NESOV, 1993
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00771.x |
|
DOI |
https://doi.org/10.5281/zenodo.10544416 |
|
persistent identifier |
https://treatment.plazi.org/id/AB4D878F-FFA3-611F-5292-F930C6B2F9A3 |
|
treatment provided by |
Marcus |
|
scientific name |
Parazhelestes robustus |
| status |
|
PARAZHELESTES ROBUSTUS NESOV, 1993
FIGURES 18–20 View Figure 18 View Figure 19 View Figure 20
(See Appendix 4 for synonymies, referred illustrations, and referred specimens.)
Holotype: CCMGE 70 View Materials /12455, left maxilla with heavily worn and eroded P4-5, M1-3.
Type locality and horizon: CBI-14, Dzharakuduk, Kyzylkum Desert, Uzbekistan. Bissekty Formation, Upper Cretaceous (middle- upper Turonian). Found in 1987.
Revised diagnosis: Differs from P. mynbulakensis in lower teeth averaging 13% larger (range equals 4 to 21%) and upper teeth averaging 10% larger (range equals 7 to 13%) and M3 narrower than M2 [37(1)].
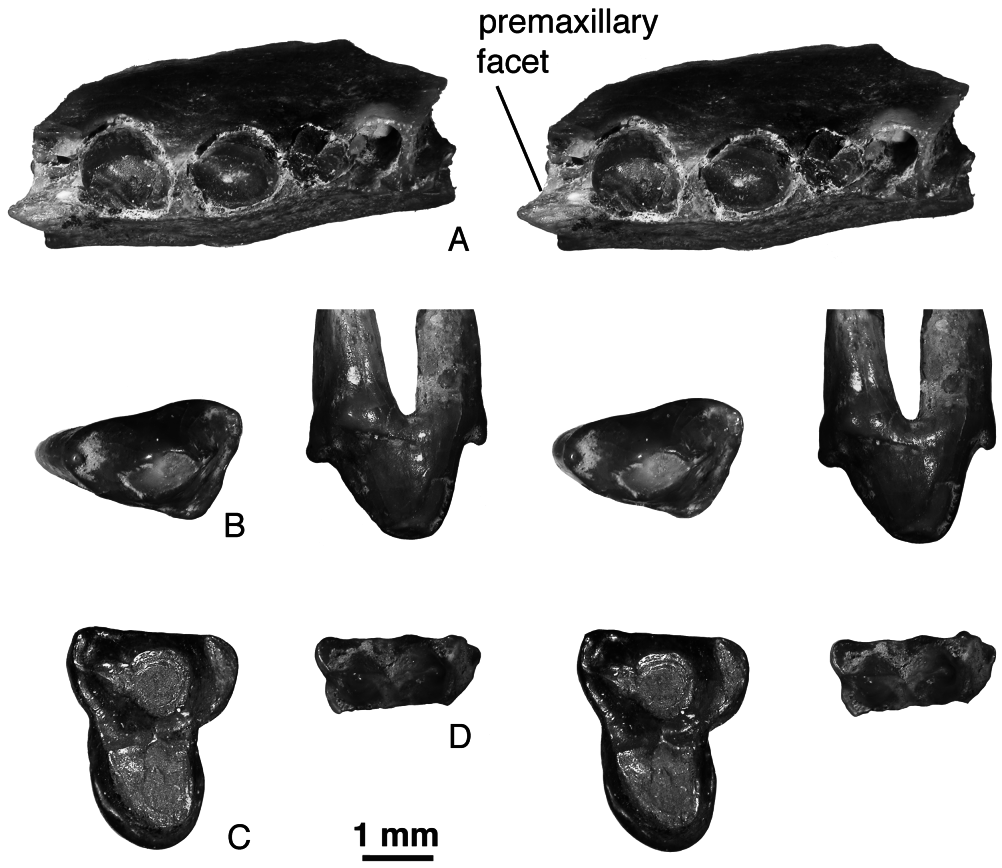
Description: Maxilla. The jugal facet on the holotype of P. robustus is higher than in P. mynbulakensis and Aspanlestes : it comes close to the alveolar border only above M3. The infraorbital foramen is slightly more posterior than in P. mynbulakensis , approximately at the level between P4 and P5. The maxilla has a distinct facet for the maxilloturbinals (holotype), as in Aspanlestes . The palatal process on URBAC 02–24 has a small premaxillary facet on the anterior end ( Fig. 18 View Figure 18 ).
Upper dentition. No upper incisors, canines, or anterior premolars are known for P. robustus . The only anterior maxillary fragment referable to P. robustus is URBAC 02–24 ( Fig. 18A View Figure 18 ). It has two approximately equal and labiolingually compressed canine roots, two roots for a small P1 set somewhat obliquely, and alveoli for a double-rooted P2 (the P2 was more than twice as long as P1). There are no diastemata between the teeth. There is some space on the maxilla anterior to the canine, but the alveolar border is broken here and it is not clear if this space contained the alveolus of the last incisor.
The P 4 is known from the holotype maxilla and an isolated specimen, URBAC 98–23 ( Fig. 18B View Figure 18 ). In the holotype the enamel of this tooth is eroded so description is based on the isolated specimen. This is a double-rooted tooth with a robust main cusp, very small mesial and distal accessory cusps, and a welldeveloped protocone swelling supported by the transversely widened distal root. There is a prominent distal ridge extending from the main cusp apex towards the posterior accessory cusp but it does not reach the latter. There is a short distolabial cingulum at the distal accessory cusp and a longer distolingual cingulum extending around the whole protocone swelling .
The P5 is known from the holotype maxilla, where the enamel of this tooth is somewhat eroded, and from isolated CCMGE 35/12176 ( Nesov, 1985a: pl. 3, fig. 1; 1997: pl. 53, fig. 5; Nesov et al., 1998: fig. 14P, Q) ( Fig. 18C View Figure 18 ). For description of CCMGE 35/12176 see Nesov et al. (1998: 59).
The DP5 labial fragment URBAC 04–270 ( Fig. 18D View Figure 18 ) is referred to P. robustus because it is larger than the DP5s of P. mynbulakensis and coincides in size with the P5 of P. robustus . The stylar shelf is of equal width (very narrow) labial to the paracone and metacone. There is a minute stylocone but no other stylar cusps.
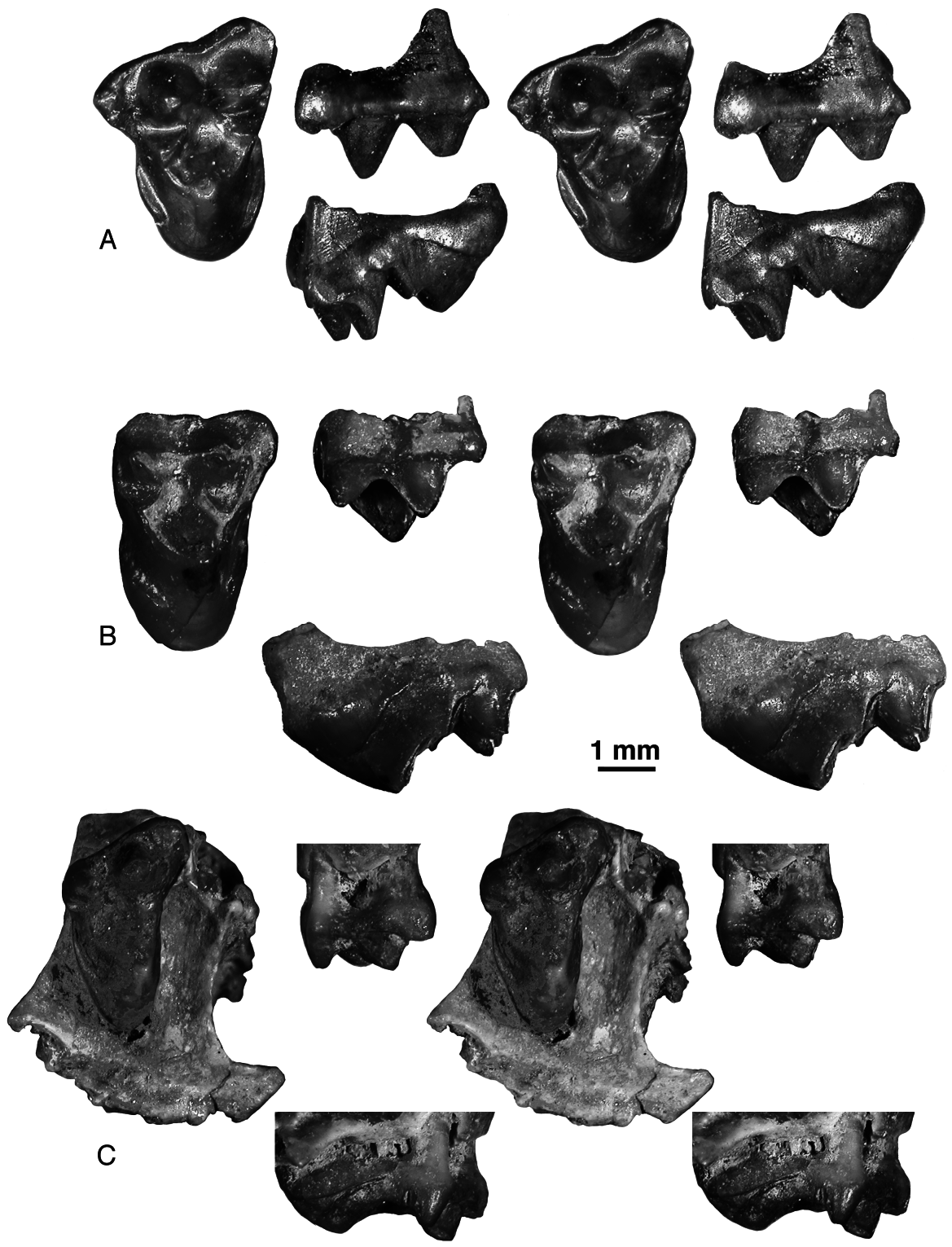
The upper molars are known from the holotype maxilla, where they are heavily worn, and from isolated specimens. The upper molars of P. robustus are larger and more robust than the upper molars of P. mynbulakensis but have basically the same structure. The main structural difference between the two species is a somewhat more labially placed protocone in P. robustus . The known sample of M1s shows almost no variation. The stylar cusp E is present in CCMGE 20/12953 ( Fig. 19A View Figure 19 ). Except for the holotype, M2 of P. robustus is known only from an isolated specimen, URBAC 99–13 ( Fig. 19B View Figure 19 ). This tooth has a relatively small parastylar lobe and the ectocingulum is expanded in the region of cusp C. The metastylar lobe was certainly small but some of the labial margin has been eroded away. The M3 is somewhat less worn in the maxilla fragment URBAC 98–22 ( Fig. 19C View Figure 19 ) compared to the holotype. Both maxillary fragments show that the lingual border of M3 is placed somewhat labial to the lingual margin of M1-2. The M 3 in URBAC 98–22 differs from M3 URBAC 03–179, referred to P. mynbulakensis , in having a relatively shorter parastylar lobe.
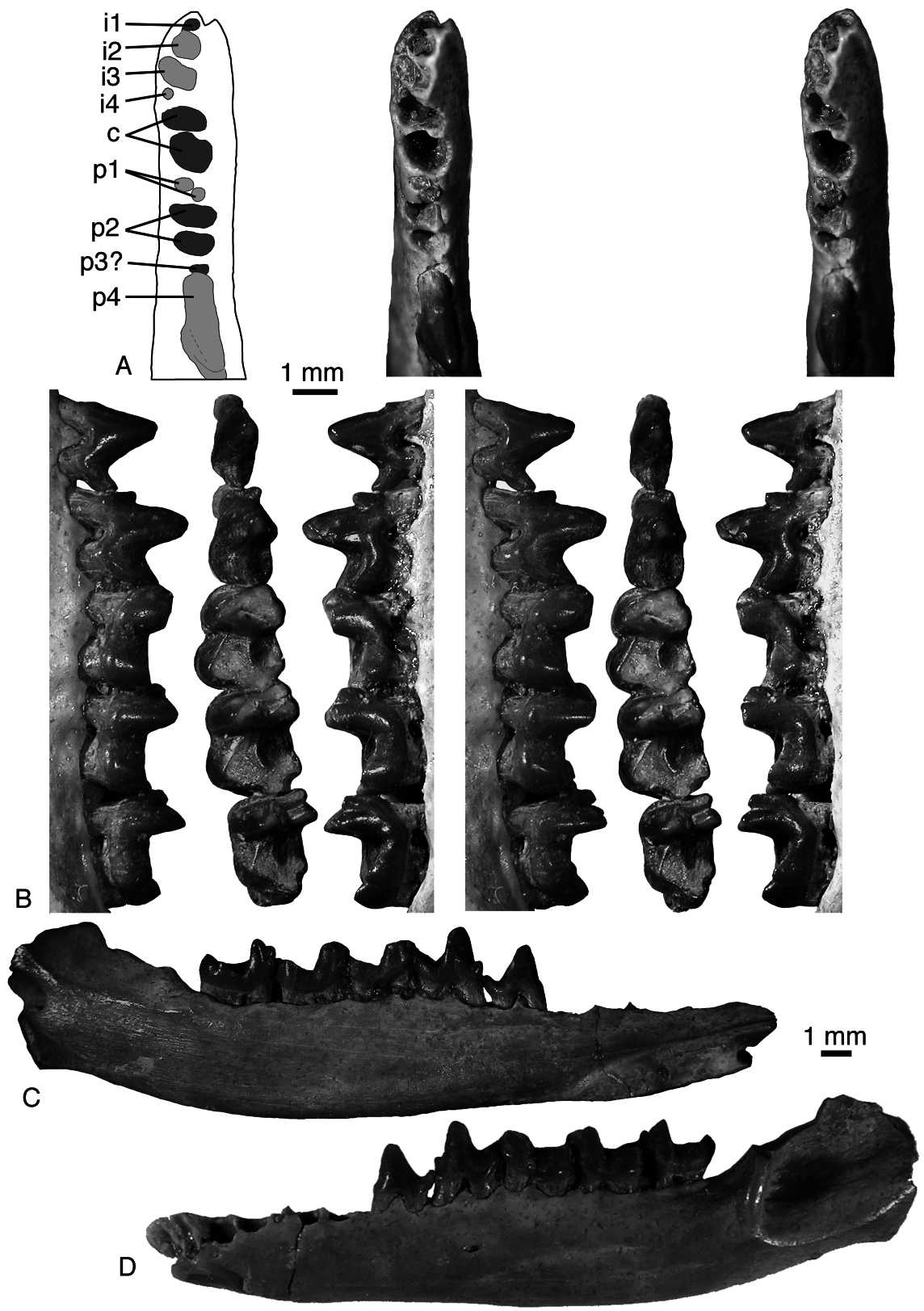
Dentary. The posterior mental foramen is usually under the distal root of p5 (ZIN 88465). In URBAC 03–120 the posterior mental foramen is very large, extending beneath the distal root of p4 and both roots of p5. The Meckelian groove is absent (URBAC 97–5, 03–120). The masseteric crest is high. There are two (URBAC 03–120, ZIN 88466) or three (ZIN 88455) labial mandibular foramina. In some specimens there is a marked tuberosity in the area of the coronoid facet of more primitive mammals, which is on the medial side of the dentary between the m3 and base of the coronoid process (ZIN 88466). In ZIN 88463 on the medial side of the coronoid process there is a robust subhorizontal crest delimiting the ventral border for the temporalis muscle. The base of the angular process and the mandibular foramen are present only in ZIN 88969; these structures do not differ from those in other zhelestids.
Lower dentition. The lower incisors, canines, and premolars (except a tentatively attributed isolated p2) are not known for P. robustus . However, in IZANUz P2155-M-5 there is a minute alveolus for i4 at the mesiolabial corner of the canine anterior root, exactly as in URBAC 99–109 of P. mynbulakensis ( Fig. 16 View Figure 16 ). This suggests that at least some individuals of P. robustus had four lower incisors as well. The lower canine is double-rooted, as evident from alveoli in IZANUz P2155-M-5. In this specimen there are no alveoli for p1 and p3, but judging from the closely spaced alveoli, it is likely to be an immature specimen and the postcanine dentition could have been dp2-4. The referral of IZANUz P2155- M-5 to the larger Eoungulatum is unlikely because the latter has a single-rooted lower canine. The alveoli for p2 and p3 are similar in size. The p3 was much smaller and mesiodistally aligned (ZIN 88489, URBAC 03–120) or oblique (ZIN 88465) to the tooth row.
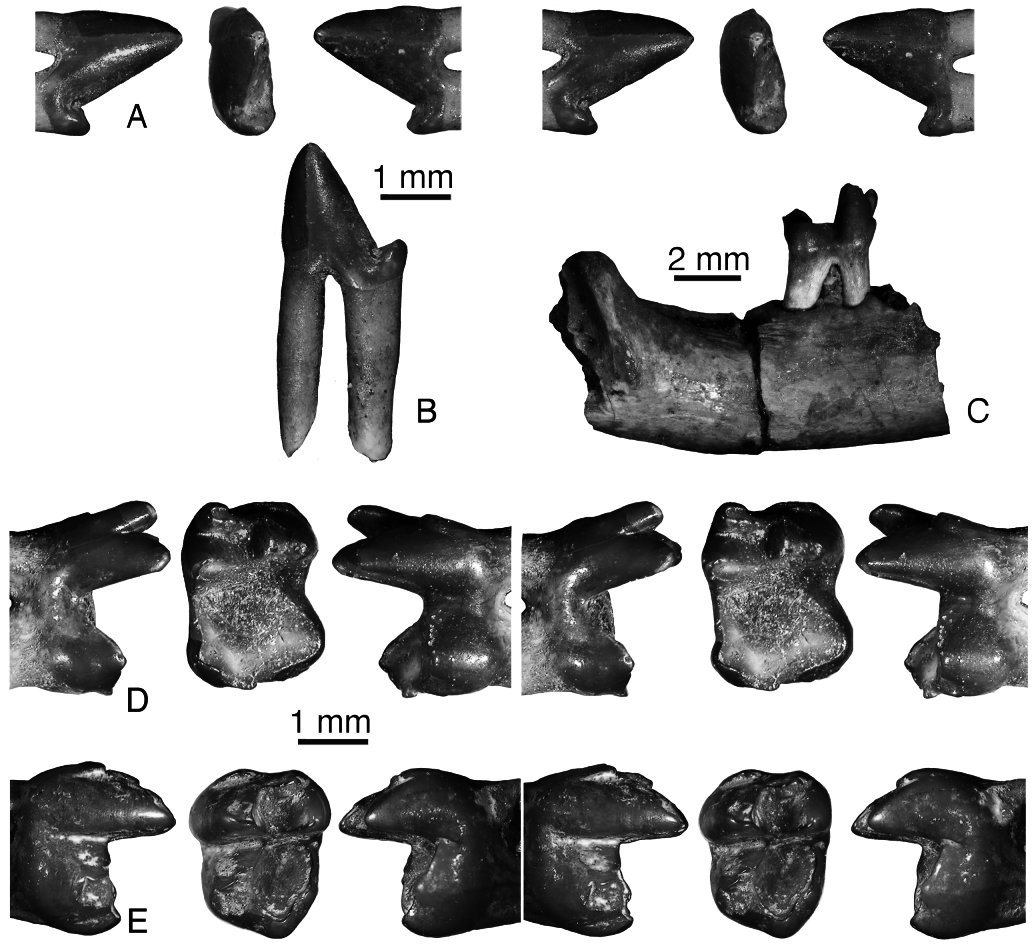
URBAC 04–324 is a lower premolar tentatively attributed to P. robustus ( Fig. 20A, B View Figure 20 ). It has two long roots, a conical central cusp, which is inclined anteriorly, no mesial accessory cusp, and a small distal accessory cusp. It is likely to be p2 because p1 and p3 are reduced in zhelestids and p4 has a distinct anterior accessory cusp. It is larger than p 2 in URBAC 97–3 and 02–13 attributed to P. mynbulakensis .
Amongst the lower molars, m1 is not known for P. robustus , m2 is known from three isolated specimens and one dentary fragment, URBAC 97–5 ( Fig. 20C, D View Figure 20 ), and m3 is known from a single isolated specimen (URBAC 03–38; Fig. 20E View Figure 20 ). The lower molars have the same morphology as in P. mynbulakensis , but are slightly larger. The postcingulid is absent in URBAC 04–260, short in URBAC 98–14, and longer in URBAC 97–5. In the latter specimen there is a faint labial cingulid. The single m3 is wider than this tooth in P. mynbulakensis , but has a much shorter talonid. The hypoconulid projects only slightly distally. The m3 paraconid is reduced and in a more labial position as in m1, whereas it is little reduced and more lingual in m2 as in P. mynbulakensis .
Measurements: See Appendices 2 and 3.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |