
Pseudopaludicola matuta, Andrade & Haga & Lyra & Carvalho & Haddad & Giaretta & Toledo, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.480 |
|
publication LSID |
lsid:zoobank.org:pub:B8FED1DD-D057-49AE-A013-12F67F8C69B9 |
|
DOI |
https://doi.org/10.5281/zenodo.3845811 |
|
persistent identifier |
https://treatment.plazi.org/id/DF080E14-2F88-45CC-876B-147D2D06C6D9 |
|
taxon LSID |
lsid:zoobank.org:act:DF080E14-2F88-45CC-876B-147D2D06C6D9 |
|
treatment provided by |
Valdenar |
|
scientific name |
Pseudopaludicola matuta |
| status |
sp. nov. |
Pseudopaludicola matuta sp. nov.
urn:lsid:zoobank.org:act:DF080E14-2F88-45CC-876B-147D2D06C6D9
Figs 2–5 View Fig View Fig View Fig View Fig , Tables 1–4 View Table 1
Pseudopaludicola sp. – Carvalho (2012): 52, Natural History Section.
Pseudopaludicola mineira from Pampulha, Belo Horizonte, Minas Gerais – Andrade et al. 2018: 87, Discussion Section.
Diagnosis
Pseudopaludicola matuta sp. nov. is assigned to the genus Pseudopaludicola by having a hypertrophied antebrachial tubercle (see Lynch 1989, Lobo 1995). The new species is characterized by the following combination of characters: (1) small size (SVL 12.8–14.2 mm in adult males); (2) absence of both, enlarged palpebral tubercles and an enlarged conical tubercle on heel; (3) relatively short hind limbs (tibiotarsal articulation only reaching the posterior margin of the eye); and (4) advertisement call composed of regular series of stereotyped two-pulsed notes, with notes having a short duration and emitted at a higher rate (notes per minute).
Differential diagnosis
Pseudopaludicola matuta sp. nov. is promptly distinguished from the species of the P. saltica group ( P. saltica , P. murundu , and P. jaredi ) by having relatively short hind limbs; i.e., the tibiotarsal articulation reaches the posterior margin of the eye in the new species, whereas it reaches the tip of the snout in species of the P. saltica group ( Andrade et al. 2016).
Pseudopaludicola matuta sp. nov. is distinguished from species of the P. pusilla group ( P. boliviana , P. ceratophyes , P. llanera , P. pusilla , and P. motorzinho ) by the absence of either T-shaped terminal phalanges or expanded toe tips (disks or pads). The tips of the phalanges of the new species are not T-shaped; they are similar in shape to those of P. falcipes (fig. 2B in Cardozo & Suárez 2012). The new species is also distinguished from P. ceratophyes by the absence of an enlarged palpebral tubercle ( Lynch 1989). Pseudopaludicola matuta sp. nov. differs from P. boliviana and P. motorzinho by the absence of an enlarged, conical tubercle on the heel ( Pansonato et al. 2016).
Pseudopaludicola matuta sp. nov. has an advertisement call composed of series of notes with two nonconcatenated pulses each ( Fig. 4 View Fig A–B); therefore, it is promptly distinguished from all species with advertisement call composed of non-pulsed notes: P. canga Giaretta & Kokubum, 2003 ( Giaretta & Kokubum 2003; Pansonato et al. 2012; Carvalho et al. 2015a), P. giarettai ( Carvalho 2012; Carvalho et al. 2015b), P. hyleaustralis Pansonato, Morais, Ávila, Kawashita-Ribeiro, Strüssmann & Martins, 2012 ( Pansonato et al. 2012), P. facureae ( Andrade & Carvalho 2013; Carvalho et al. 2015a), and P. parnaiba Roberto, Cardozo & Ávila, 2013 ( Roberto et al. 2013; Carvalho et al. 2015a); and from all species with notes with concatenated pulses (= lack of interpulse interval; sensu Magalhães et al. 2014): P. mystacalis [12–14 concatenated pulses; Pansonato et al. 2013], P. boliviana [3–6; Duré et al. 2004], P. ibisoroca Pansonato, Veiga-Menoncello, Mudrek, Jansen, Recco-Pimentel, Martins & Strüssmann, 2016 [3–12; Pansonato et al. 2016], and P. motorzinho [2–6; Pansonato et al. 2016].
Pseudopaludicola matuta sp. nov. is distinguished from other congeners with notes with non-concatenated pulses (values within square brackets “[]”) by the following acoustic traits: P. ameghini (Cope, 1887) has longer note duration (27–52 [60–121] ms), higher number of pulses per note (2 [3–5]), and lower note rate (542–1101 [348–452] notes per minute) ( Andrade et al. 2017a); P. ternetzi Miranda-Ribeiro, 1937 has higher number of pulses per note [3–6] ( Andrade et al. 2017a); P. atragula Pansonato, Mudrek, Veiga-Menoncello, Rossa-Feres, Martins & Strüssmann, 2014 has longer note duration [300–700 ms], higher number of pulses per note [9–36], and lower note rate [42–98 notes per minute] ( Pansonato et al. 2014). The three species of the P. saltica species group have lower note rate [180–480 notes per minute, combined values], and vary highly the number of pulses in their notes (2–7 pulses per note, combined values; Andrade et al. 2016). Pseudopaludicola falcipes and P. restinga Cardozo, Baldo, Pupin, Gasparini & Haddad, 2018 have lower note rates [238–535 notes per minute] ( Andrade et al. 2018; Cardozo et al. 2018). Pseudopaludicola pocoto Magalhães, Loebmann, Kokubum, Haddad & Garda, 2014 and P. florencei Andrade, Haga, Lyra, Leite, Kwet, Haddad, Toledo & Giaretta, 2018 have stereotyped three-pulsed notes along their series of notes, longer note duration [108–397 ms, combined values], and lower note rate [100–297 notes per minute, combined values] ( Magalhães et al. 2014; Andrade et al. 2017b, 2018). In comparison with the phylogenetically close related species, the new species is distinguished from P. mineira by having shorter note duration [50–114 ms] and lower note rate [282–485 notes per minute] ( Table 2). The advertisement call of P. ceratophyes , P. llanera , and P. pusilla are unknown; however, there are strong morphological differences among them and P. matuta sp. nov.
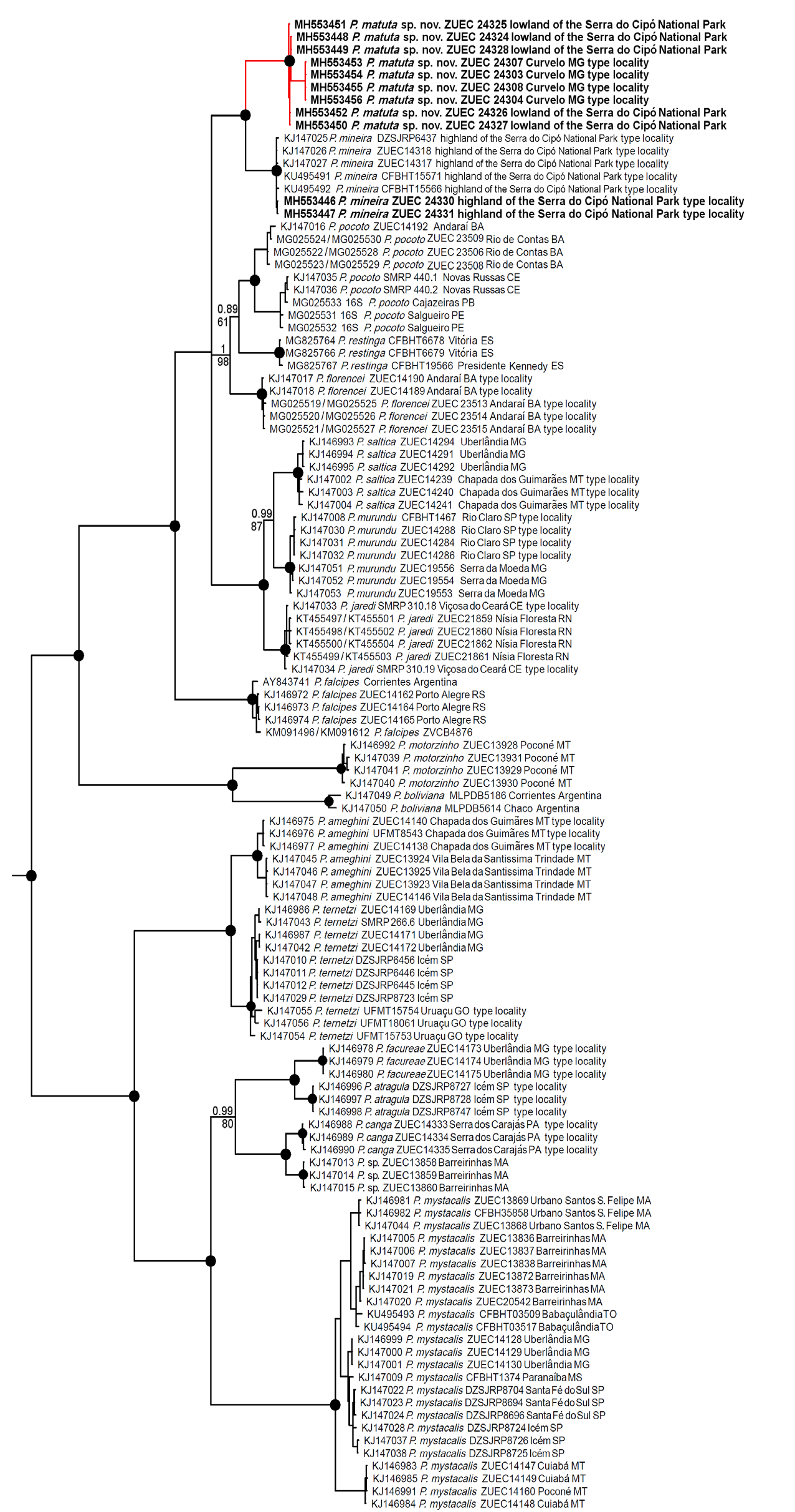
In relation to the most closely related species, P. mineira , the randomForest multivariate approach applied to morphometric data indicated a broad overlap between the two partitions ( Fig. 5 View Fig A–B), with a considerable classification error ( Table 3). We found no statistical significance in any trait. Therefore, we were unable to distinguish P. matuta sp. nov. from P. mineira based on morphometric or any other feature of external morphology or coloration. In contrast, the RandomForest multivariate approach on acoustic data highlighted a full segregation between P. matuta sp. nov. and P. mineira ( Table 3, Fig. 5C View Fig ), without any classification error. Pulse rate, note duration, notes per minute, and interpulse interval are the main sources of variation in both variable importance measurements ( Fig. 5D View Fig ). The trait of notes per minute for P. mineira did not overlap the values for the new species either. It is noteworthy that the air temperature overlapped at the time of the field recordings for both species ( Table 2), and is among the variables that least explained the variation of the dataset, with low importance measures as produced by randomForest ( Fig. 5D View Fig ). Therefore, we do not attribute this found acoustic difference to the influence of the air temperature. In addition, there was only a slight overlap between both species in note duration (see Table 2). The new species can be significantly differentiated from P. mineira in the following traits: note duration, internote interval, interpulse interval, pulse rate, and dominant frequency (for all these traits, Wilcoxon-Mann-Whitney Tests had P <0.01).
Etymology
The feminine noun matuta in Brazilian Portuguese means rustic, provincial, related to those who live in the countryside.
Type material
Holotype
BRAZIL: adult Ƌ, Minas Gerais, municipality of Curvelo, Sítio Mato do Engenho , 18°46′07.6″ S, 44°26′50.7″ W, 620 m a.s.l., 29 Nov. 2017, F.S. Andrade & I.A. Haga leg. ( ZUEC 24302 View Materials ; Figs 2–3 View Fig View Fig , call voucher).
GoogleMapsParatopotypes
BRAZIL: 12 adult ƋƋ, same collection data as the holotype ( ZUEC 24303–6, 24308–10, 24313, 24315– 8); 3 adult ƋƋ, same locality as holotype, 21 Feb. 2011, T. R. Carvalho leg. ( AAG-UFU 0308, 0386–7); 5 adult ♀♀, same collection data as the holotype ( ZUEC 24307, 24311–2, 24314, 24323).
Type locality BRAZIL: Minas Gerais, municipality of Curvelo, Sítio Mato do Engenho, 18°46′07.6″ S, 44°26′50.7″
W, 620 m a.s.l.
Other material examined
BRAZIL: 3 adult ƋƋ, Minas Gerais, municipality of Santana do Riacho, district of Serra do Cipó, southern foothills of Serra do Cipó National Park, near Lagoa da Capivara (19°20’46.30” S, 43°36’59.62” W, 797 m a.s.l.), 2–3 Dec. 2017, F.S. Andrade & I.A. Haga leg. ( ZUEC 24324, 24327, 24329); 3 adult ♀♀, same collection data as previous ( ZUEC 24325–6, 24328).
Description of the holotype
Body elliptic and broad ( Table 1 View Table 1 , Fig. 2 View Fig A–B). Head elliptical, slightly wider than long. Snout subovoid in dorsal view and rounded in profile ( Fig. 2 View Fig C–D). Eye not protuberant. Eye diameter almost equal to the interorbital distance. Interorbital area flat. Pupil rounded. Upper eyelid without tubercles. Nostril not protuberant and closer to the snout tip than to the eye. Canthus rostralis rounded, smooth. Loreal region slightly concave. Single subgular vocal sac, externally expanded and with discrete longitudinal folds. Choanae rounded, well separated from each other. Vocal slits present. Tympanum indistinct. A discrete tympanic ridge from behind the eye to the proximal portion of the arm. Mouth opening ventral. Vomerine teeth absent. Tongue elliptical, longer than wide, posteriorly free, without pigmentation at its base. Flank with discrete granules. One ovoid antebrachial tubercle present in the first quarter of the forearm and a second ovoid tubercle closer to elbow. Finger and toe tips not expanded. Outer and inner metacarpal tubercles well-defined, outer metacarpal tubercle ovoid and inner metacarpal tubercle rounded. Fingers with single and rounded subarticular tubercles. Supernumerary tubercles absent on palm of hand. Thumb with a keratinized, light brown nuptial pad, extending from the base of the hand to the proximal limit of the terminal phalanx, covering almost the entire external portion of the finger. Webbing absent between fingers. Relative finger lengths, when adpressed one to another: I <II <IV <III ( Fig. 2E View Fig ). Outer metatarsal tubercle well defined, conical. Inner metatarsal tubercle elliptical. The internal metatarsal tubercle larger than the external. Toes with well-defined, single, enlarged, and rounded subarticular tubercles. Supernumerary tubercles absent on sole of the foot. Toes webbed basally and fringed along their sides to almost their tips. Fringes developed on all toes (mainly II, III, IV, and V). External fringe on Toe V continues almost to the outer metatarsal tubercle. Well-developed fold from internal metatarsal tubercle to the mid-ventral tarsus, ending in a tarsal tubercle poor protuberant. Relative toe lengths, when adpressed one to another: I <II <V <III <IV ( Fig. 2F View Fig ). Hind limb robust and moderately long with the tibiotarsal articulation just reaching the posterior margins of eye. Thigh shorter than tibia. Foot longer than thigh. Foot longer than tibia. Tubercle absent on heel. Belly skin smooth. Abdominal fold present and complete. Dorsal surfaces of head, body, and limbs smooth. Cloacal region smooth ( Fig. 2B View Fig ). Measurements of the holotype presented in Table 1 View Table 1 .
Color pattern of the holotype in preservative
Dorsum grayish with dark gray, white, and brown blotches. Belly whitish (unpigmented). Throat light beige, unpigmented. Throat darker than belly. Dorsum darker than the dorsal surfaces of limbs. Region between upper lip and eye with several rounded white blotches. Ventral faces of arms and legs unpigmented, except that of the thigh (slight pigmented). Palm of hand pigmented. Sole of foot pigmented and darker than hands, arms, and hind limbs. Color of the sole of the foot similar to that of the dorsum of hind limb. Dorsal faces of arms light brown with several dark brown blotches. Dorsal faces of legs light grayish with scattered brown blotches. Dark brown nuptial pads ( Fig. 2 View Fig ).
Variation in the type series
Dorsal surface of body varies from dark gray to dark brown, with black or dark brown irregular blotches ( Fig. 3 View Fig ). The specimens ZUEC 24303–18 and 24323 have transverse stripes on thighs, shanks, and feet. The specimens ZUEC 24304, 24310–3, 24317–8 have a well-defined light vertebral line. The specimens ZUEC 24304, 24310–3, 24315, 24317–8, and 24323 have dorsolateral stains on body, from posterior corner of eyes to almost the region of insertion of legs. The specimens ZUEC 24304–5, 24307–8, 24311, 24315–8, 24323 also have white blotches on the region between the posterior corner of eyes and the region of insertion of arms. The specimen ZUEC 24312 has the region between upper lip and eye with alternating vertical gray and light beige stripes. Females have a more robust body and lack nuptial pads, vocal sac, and vocal slits.
Advertisement call
Pseudopaludicola matuta sp. nov. emits the advertisement call with highly variable duration (3.6– 66.2 s), consisting of series of stereotyped two-pulsed notes (1–9 series of two-pulsed notes per call) that lasts 0.2– 34.0 s, separated by intervals of 0.2– 1.9 s ( Fig. 4A View Fig ). Before the emission of the series of stereotyped two-pulsed notes, 1–10 (mean = 4.4, SD = 3.3) isolated pulsed notes with irregular structure, duration, interval, and number of pulses are emitted, herein referred to as introductory notes ( Fig. 4A View Fig ). Introductory notes last 29–117 ms (mean 64, SD = 24), separated by intervals of 37–495 ms (mean = 213, SD = 111), and are composed of 2–5 non-concatenated pulses (mean = 3.3, SD = 0.7). These pulses vary from 4–16 ms (mean = 9, SD = 2), separated by intervals of 1–36 ms (mean = 16, SD = 6). Dominant frequency peaks are between 4.1–4.5 kHz (mean = 4.4, SD = 0.2). On the other hand, within the series of stereotyped two-pulsed notes, the notes have regular structure, duration, interval, and number of pulses. These notes last 27–52 ms, separated by intervals of 17–76 ms, and are released at a rate of 542–1,101 notes per minute; notes have a slightly increase in amplitude from the first to the second pulse (see oscillogram in Fig. 4B View Fig ). Pulses vary from 4–15 ms, separated by intervals of 11–40 ms, and are released at a rate of 43–74 pulses per second ( Fig. 2B View Fig ). Dominant (= fundamental) frequency peaks are between 3.9–5.1 kHz; the minimum frequency ranges between 1.8–4.6 kHz and the maximum frequency ranges from 4.4–5.5 kHz. Notes have up to three harmonics; the second ranging from 6.9–9.8 kHz (mean = 8.7; SD = 0.4). Air temperature of recorded calls varied from 24.0 to 25.2°C. Call quantitative traits are summarized in Table 2.
Traits that were classified as static (within-male CV <5%) to P. matuta sp. nov. were note duration (mean = 4.2%, SD = 1.8 [1.6–7.4]), number of pulses per note (mean = 0%), pulse rate (mean = 4.2%, SD = 1.8 [1.6–7.4]), and dominant peak (mean = 0.9%, SD = 0.7 [0.0–3.0]). The other traits were classified as dynamic (CV <7.4%). Additionally, the between-male coefficient of notes per minute was equal to 9.2%, this low variability among males allowed us to classify this trait as static.
We noticed a remarkable intraspecific variation in the advertisement call of P. matuta sp. nov., in which the three males of the population from the southern foothills of the Serra do Cipó National Park have a lower note rate (529–737 notes per minute) than those of male types (738–1101 notes per minute). It is noteworthy that the air temperatures measured during the recordings in these two localities did not overlap, 24.0–25.2°C at type locality, and 20.5–23.0°C in lowlands of the Serra do Cipó National Park.
Phylogenetic inferences and mitochondrial DNA divergences
The inferred tree topology agreed with previous phylogenetic analyses of Pseudopaludicola (Veiga- Menoncello et al. 2014; Andrade et al. 2016, 2018), and was very similar between BI and ML inferences ( Fig. 6 View Fig ). The new species was found as sister clade of P. mineira ( Fig. 6 View Fig ). Uncorrected genetic distance between the new species and P. mineira was 3.5% (mean value) and maximum intraspecific distance was 0.1% ( Table 4). No molecular data are available for P. ceratophyes , P. llanera , P. pusilla , P. hyleaustralis , P. parnaiba , P. giarettai , P. ibisoroca ; however, the new species is strongly diagnosed from these species by morphology and acoustics (see further details in Differential diagnosis section).
Natural history notes
We recorded specimens of P. matuta sp. nov. in an area with relatively dense vegetation. In this site, there were grasses, shrub vegetation, and some spaced trees of 5–7 m in height, with the soil well soaked. Males of other Pseudopaludicola species usually call exposed in open areas. Curiously, two other specimens of P. matuta sp. nov. were recorded a few meters from this above-mentioned site in an open and waterlogged area, where P. giarettai also occurs syntopically. In both places, we observed dozens of specimens of P. matuta sp. nov.
However, during the fieldwork in the lowlands of the Serra do Cipó National Park, we were able to find a single small population near Lagoa da Capivara and Cipó River. During the two sampling nights, it was possible to hear only eight males, which were recorded. They were not excited and were vocalizing wellspaced from each other. This social condition may have influenced the intraspecific acoustic variation described by us. Besides that, air temperature was lower (without overlap) during the fieldwork in the lowlands of the Serra do Cipó National Park.
Distribution
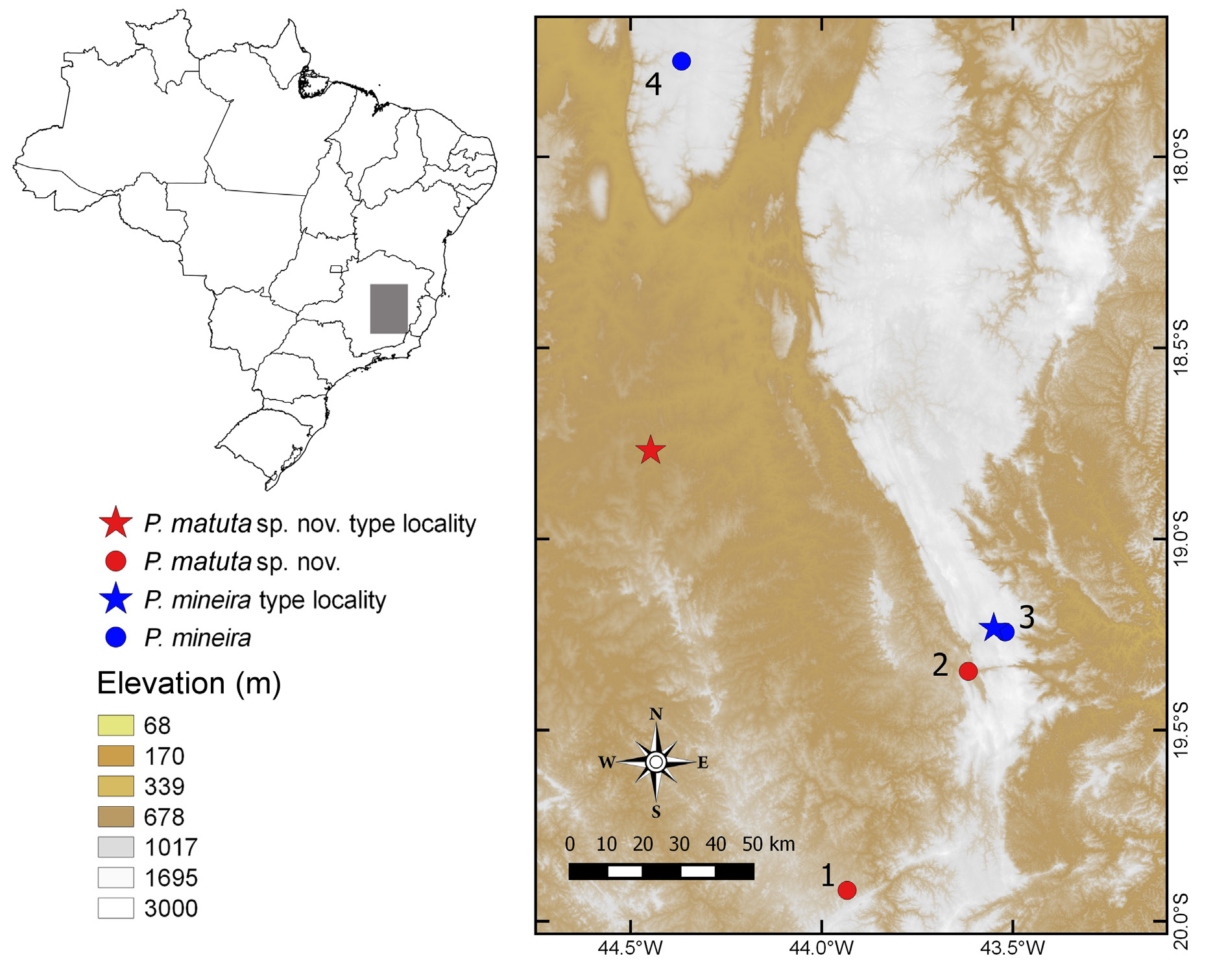
Pseudopaludicola matuta sp. nov. is known from its type locality, lowlands of the Serra do Cipó National Park (southern foothills), district of Serra do Cipó, municipality of Santana do Riacho, and in the municipality of Belo Horizonte from a report in the 1960s ( Fig. 1 View Fig ). However, it is not possible to find this species nowadays in the municipality of Belo Horizonte (F.S.F. Leite, pers. obs.). Lobo (1994) reported the occurrence of P. mineira in the Serra do Cipó National Park (at 1264 m a.s.l., type locality) and in Serra do Cabral (at 1104 m a.s.l.), a nearby mountainous area but isolated from the main Espinhaço mountain range ( Fig. 1 View Fig ). The altitudes we found P. matuta sp. nov. vary between 620 and 883 m a.s.l. In addition, in the type locality and where we collected P. mineira during our fieldwork are only about 14 km northeast from the site where we recorded P. matuta sp. nov. (797 m a.s.l., Fig. 1 View Fig ); however, the elevational difference between these two sites is about 460 m. We did not find P. matuta sp. nov. during our fieldwork in the rupestrian grasslands of the Serra do Cipó National Park. Therefore, it seems that P. mineira is restricted to the higher rupestrian grasslands of the Serra do Cipó National Park and Serra do Cabral ( Lobo 1994), whereas P. matuta sp. nov. occurs in adjacent lowland regions to the west and south of this unique Brazilian mountain range.
The advertisement call and acoustic diagnosis of P. mineira
Pseudopaludicola mineira also emits long advertisement calls, consisting of series of stereotyped twopulsed notes that lasts 0.4– 44.3 s, separated by intervals of 0.3– 10.9 s ( Fig. 4C View Fig ). Notes last 50–114 ms separated by intervals of 42–159 ms and released at a rate of 282–485 notes per minute. Only six notes of one male have three non-concatenated pulses of all the analysed notes (n = 180 notes); the other notes have two non-concatenated pulses each. Pulses vary from 4–22 ms, are separated by intervals of 5–94 ms, and are released at a rate of 17–118 pulses per second ( Fig. 4D View Fig ). Dominant (= fundamental) frequency peaks between 3.6–4.3 kHz, the minimum frequency ranges 2.3–3.9 kHz, and the maximum frequency ranges 4.0–4.5 kHz. Second harmonic peaks between 6.8–8.1 kHz (mean = 7.1; SD = 0.07). Pseudopaludicola mineira also has introductory notes with irregular structure, duration, interval, and number of pulses. Quantitative traits are summarized in Table 2. Air temperature of recorded calls varied from 16.0 to 25.5°C.
Similarly to the new species, P. mineira can be distinguished from species that have non-pulsed structure or with concatenated pulses (with lack of interpulse interval): P. canga , P. giarettai , P. hyleaustralis , P. facureae , P. parnaiba , P. mystacalis , P. boliviana , P. ibisoroca , and P. motorzinho . Pseudopaludicola mineira distinguishes from other congeners [values within square brackets] by the following acoustic traits: P. ternetzi has higher note rate (282–485 [606–921] notes per minute), and higher number of pulses per note (2–3 [3–6]) ( Andrade et al. 2017a); P. ameghini has higher number of pulses per note [3–5] ( Andrade et al. 2017a); P. atragula has longer note duration (50–144 [300–700 ms], and higher number of pulses per note [9–36], and lower note rate [42–98 notes per minute] ( Pansonato et al. 2014). From the three long-legged species ( P. saltica , P. murundu , and P. jaredi ), P. mineira is distinguished by having low variation in the number of pulses per note [2–7] ( Andrade et al. 2016). Pseudopaludicola pocoto and P. florencei have lower note rate [100–297 notes per minute, combined values], higher dominant frequency (3.6–4.3 [4.2–6.5] kHz), and stereotyped three-pulsed notes ( Magalhães et al. 2014; Andrade et al. 2017b, 2018). Pseudopaludicola mineira differs from P. falcipes by its lower dominant frequency [4.7–6.0 kHz; Andrade et al. 2018; present study]; however, the advertisement calls of these two species are quite similar in the temporal domain. We were unable to distinguish acoustically P. mineira from P. restinga based on data from literature ( Cardozo et al. 2018). Note emission rate is one of the most informative acoustic traits to diagnose closely related species of Pseudopaludicola (e.g., Andrade et al. 2017a, 2018). In Cardoso et al. (2018) there is no measure of this feature but an estimated value from their figured sounds is around 490 notes per minute, what is within the range we observed to P. mineira .




| ZUEC |
Museu de Zoologia da Universidade Estadual de Campinas |
| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pseudopaludicola matuta
| Andrade, Felipe Silva de, Haga, Isabelle Aquemi, Lyra, Mariana Lúcio, Carvalho, Thiago Ribeiro de, Haddad, Célio Fernando Baptista, Giaretta, Ariovaldo Antonio & Toledo, Luís Felipe 2018 |
Pseudopaludicola mineira
| Andrade F. S. & Haga I. A. & Lyra M. L. & Leite F. S. F. & Kwet A. & Haddad C. F. B. & Toledo L. F. & Giaretta A. A. 2018: 87 |
Pseudopaludicola
| Carvalho T. R. 2012: 52 |