
Australotarsius Solodovnikov & Newton
|
publication ID |
https://doi.org/ 10.5281/zenodo.186314 |
|
DOI |
https://doi.org/10.5281/zenodo.6226722 |
|
persistent identifier |
https://treatment.plazi.org/id/A641F03D-FFBC-6705-FF5F-F951FA966072 |
|
treatment provided by |
Plazi |
|
scientific name |
Australotarsius Solodovnikov & Newton |
| status |
|
Australotarsius Solodovnikov & Newton View in CoL , new genus
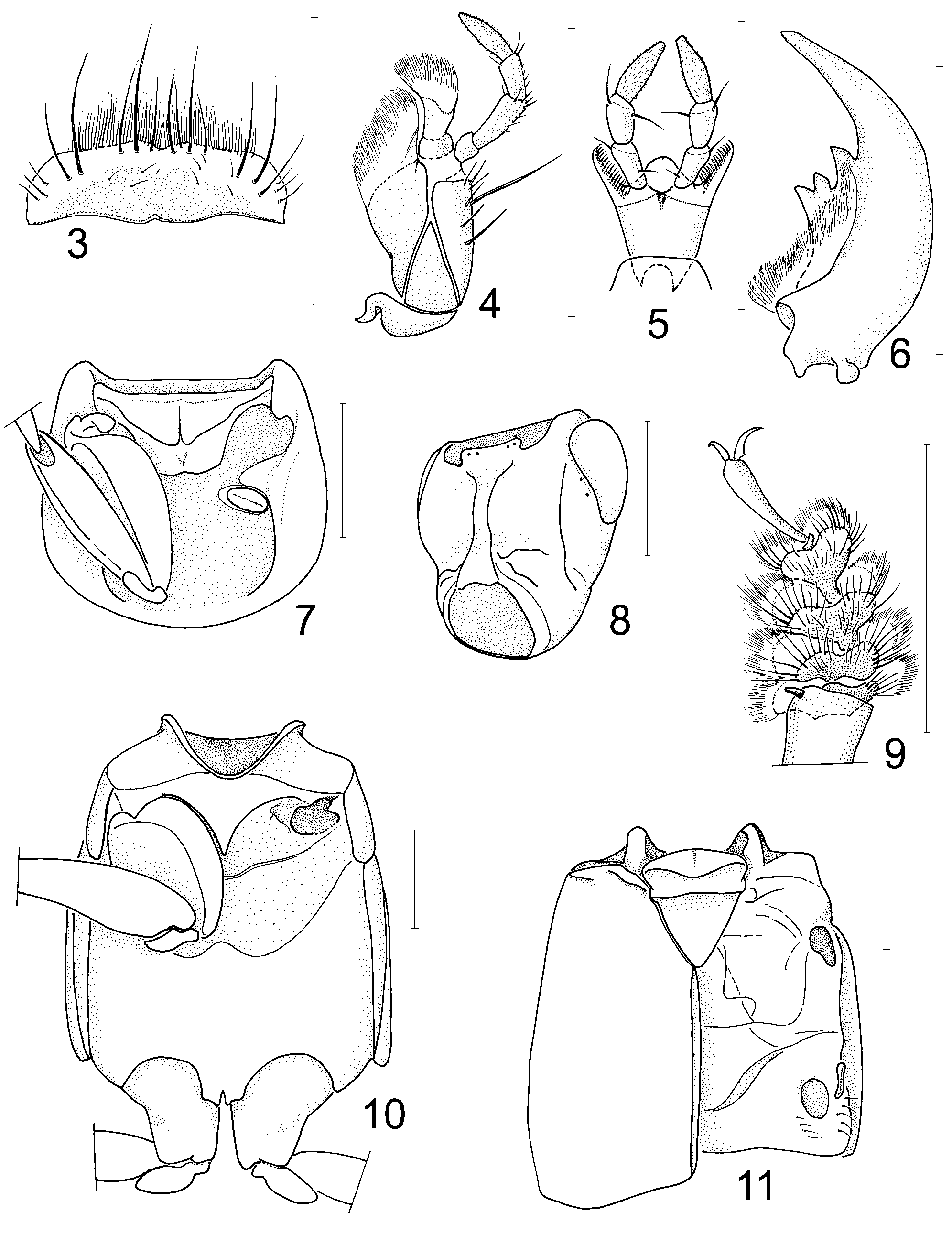
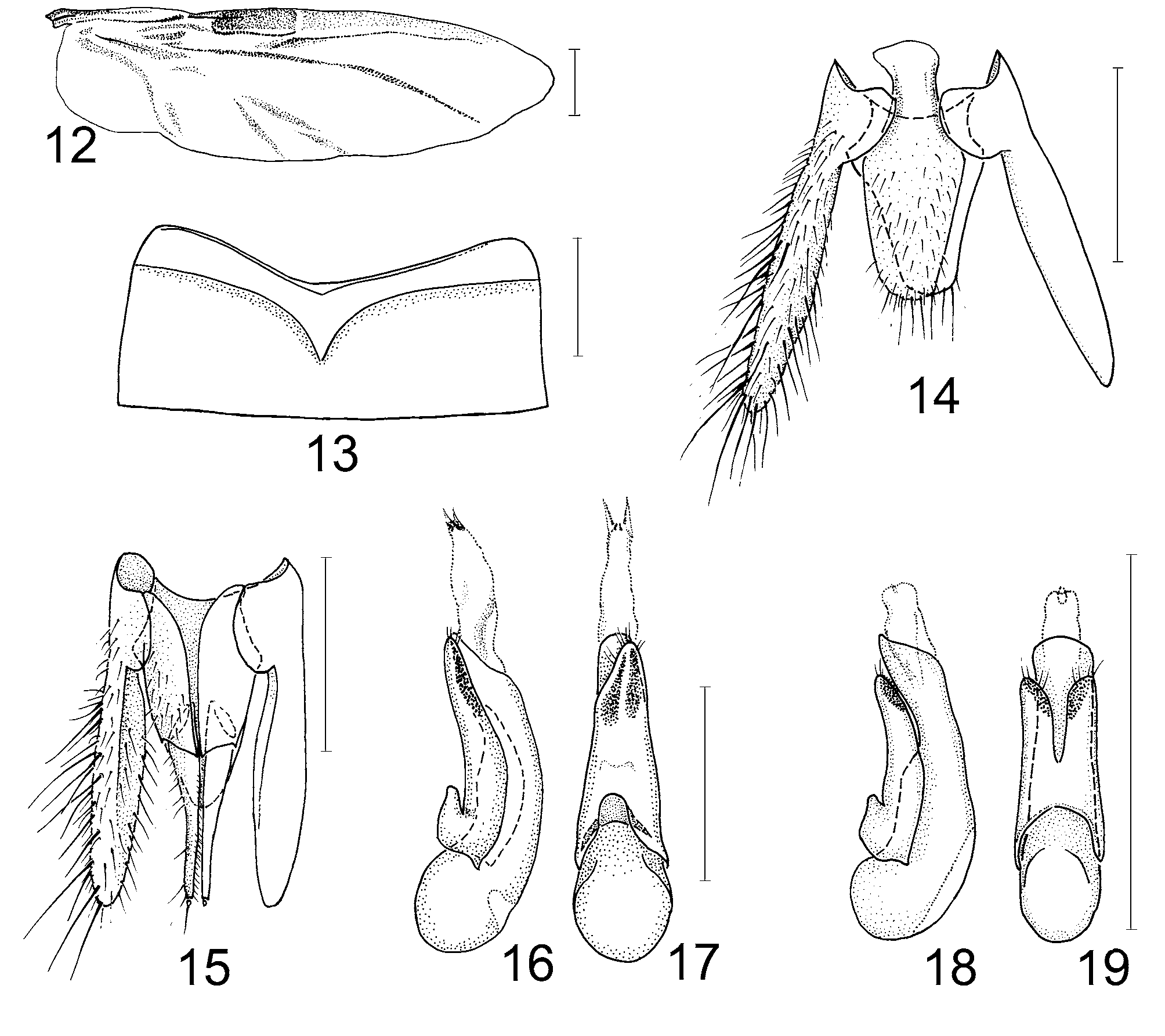
( Figs. 1–19 View FIGURES 1 – 2 View FIGURES 3 – 11 View FIGURES 12 – 19 )
Type species. Australotarsius grandis Solodovnikov and Newton , new species.
Diagnosis. From all other genera of Staphylinini , Australotarsius can be distinguished by the following combination of characters: entire body evenly setose; apical segment of maxillary palpus setose; neck broad; infraorbital ridges well developed; anterior tarsi in both sexes very broad; male sternite VIII without secondary sexual modifications, of the same shape as in females.
Description. Habitus as in Figs 1, 2 View FIGURES 1 – 2 . Entire body covered by dense setation. Body length 7.4–14 mm.
Head capsule ( Figs 1–2 View FIGURES 1 – 2 , 8 View FIGURES 3 – 11 ) rounded with well developed, moderately large eyes; nuchal constriction indicated only laterally (in A. grandis ) or laterally and dorsally (in A. tasmanicus ), in the former case nuchal ridge developed only laterally, in the latter—laterally and dorsally; epistomal suture absent; infraorbital ridges present, extending to about the middle of the distance from neck to base of mandibles; postgenal and ventral basal ridges well developed; postmandibular ridge and dorsal basal ridge absent. Antennal insertions at anterolateral margins of frons, anterior to eyes, not concealed from above; distance between them slightly longer than distance from either insertion to margin of eye. Antennae moderately long, with distinctly elongate first antennomere (as long as second and third antennomeres together); first three antennomeres setose, but without pubescence, remaining antennomeres setose and pubescent. Mandibles ( Fig. 6 View FIGURES 3 – 11 ) moderately long, symmetrical, each with teeth on inner side, without dorso-lateral mandibular groove; prostheca broadly attached near base of mandible, mola not developed. Labrum ( Fig. 3 View FIGURES 3 – 11 ) strongly transverse, without transparent apical membrane, only slightly notched medio-apically. Maxilla ( Fig. 4 View FIGURES 3 – 11 ) with lacinia and galea densely hairy at apex; stipes with clear sutures; maxillary palpus four-segmented, last segment fusiform, finely setose. Labium ( Fig. 5 View FIGURES 3 – 11 ): ligula small, only slightly notched apically, paraglossae long with comb of stout setae internally; last segment of labial palpus fusiform with slightly truncate apex, finely setose.
Pronotum ( Fig. 7 View FIGURES 3 – 11 ) with anterior angles projecting anteriad over apical margin of prosternum; pronotal hypomera slightly inflected inwards, only slightly visible in lateral view; superior marginal line of pronotum well developed throughout its whole length, not deflexed ventrad; inferior marginal line shorter, not meeting with superior marginal line; pronotal hypomeron with weakly sclerotized postcoxal process. Pronotosternal sutures visible but not membranous. Prosternum with midlongitudinal carina, without large conspicuous macrosetae. Mesoventrite without large conspicuous macrosetae, with narrow, acute mesoventral intercoxal process. Meso-metaventral suture well developed, membranous. Mesoscutellum ( Fig. 11 View FIGURES 3 – 11 ) with two transverse carinae: one very close to base, another close to middle. Elytra ( Fig. 11 View FIGURES 3 – 11 ) relatively long, with sharp humeri and well developed sub-basal ridge immediately adjacent to elytral articulation and extending anteriad to anterior margin of elytron; epipleural part gradually deflexed, lacking epipleural ridge; sutural angle distinct. Hind wings ( Fig. 12 View FIGURES 12 – 19 ) fully developed, with veins CuA and MP4 completely separate. Metaventrite ( Fig. 10 View FIGURES 3 – 11 ) well developed, with deep mesocoxal acetabuli delimited by carina. Legs moderately long and slender; anterior and middle (smaller) coxae large, conical, contiguous ( Figs. 7, 10 View FIGURES 3 – 11 ). Hind coxae ( Fig. 10 View FIGURES 3 – 11 ) as long as wide, almost contiguous at base. Anterior tibia densely setose, without spines externally; middle and posterior tibiae setose, with spines externally. Tarsal formula 5-5-5; claws moderately long, arcuate; pair of empodial setae on each tarsus, shorter on anterior tarsi and very long, longer than claws on middle and posterior tarsi. Anterior tarsi ( Fig. 9 View FIGURES 3 – 11 ) in both sexes with tarsomeres I–IV strongly dilated and with dense, long whitish adhesive setae ventrally.
Abdomen more or less parallel-sided along most of its length, narrowed posteriad, slightly flattened dorsoventrally; abdominal tergite I with protergal glands externally manifested by deep indentation (acetabulum) at each side of tergite and adjacent groups of setae ( Fig. 11 View FIGURES 3 – 11 ). Segments III–VI each with two pairs of elongate paratergites, segment VII with two pairs of paratergites, basal short one and apical long one; segment VIII without paratergites; tergites III–VII only with one basal carina; sternite III with basal carina medially strongly angulate ( Fig. 13 View FIGURES 12 – 19 ). Sternite VIII without apical emargination in males, with apical margin similarly rounded in both sexes. Abdominal intersegmental membranes attached preapically and having a pattern of small irregular rounded sclerites. Male ( Fig. 14 View FIGURES 12 – 19 ) and female ( Fig. 15 View FIGURES 12 – 19 ) tergite IX consisting of two elongate, inflated lateral sclerites, apically obtuse, widely separated dorsally by tergite X and ventrally by sternite IX in males or valvifers (first gonocoxites) in females. Male sternite IX ( Fig. 14 View FIGURES 12 – 19 ) elongate, more or less symmetrical, with short asymmetrical basal portion.
Aedeagus ( Figs. 16–19 View FIGURES 12 – 19 ): median lobe elongate, apically obtuse, symmetrical; paramere with peg setae on the underside, and four pairs of setae apically. Aedeagus rotated by 90° in the abdomen in repose: its dorsal (parameral) side facing left laterally.
Female external genitalia consisting of a pair of large basal valvifers, short coxites (second gonocoxites) and tiny styli; female tergite X symmetrical, large ( Fig. 15 View FIGURES 12 – 19 ).
Etymology. The genus name is derived from two words “ Australia ” and “tarsus”. It refers to the country of origin and spectacular wide anterior tarsi of this new taxon. It is Latinized noun of masculine gender.
Comparison and phylogenetic relationships. Due to somewhat inflexed hypomera of pronotum and well developed infraorbital ridges Australotarsius looks like a member of the conventional subtribe Quediina (for its definition and list of included taxa see, for example, Smetana & Davies 2000 and Herman 2001, respectively), which has been shown to be a polyphyletic group ( Solodovnikov 2006; Solodovnikov & Schomann 2009). Within the newly emerging phylogenetic framework for Staphylinini , Australotarsius can be placed in the clade “Quediina propria”, which, according to Solodovnikov and Schomann (2009, Fig. 1 View FIGURES 1 – 2 ), includes two subclades: one with the north temperate members of the genus Quedius Stephens, 1829 and the genus Indoquedius Blackwelder, 1952 and another with the genera Anchocerus Fauvel, 1905 , Acylophorus Nordmann, 1837 , Euryporus Erichson, 1839 and Hemiquedius Casey, 1915 . Presumably Australotarsius belongs to that latter subclade of “Quediina propria”. The exact limits of this lineage within Staphylinini are not finally clear yet: apparently it also includes the genus Anaquedius Casey, 1915 , but may not include Euryporus . All these genera here affiliated with Australotarsius share numerous character states (e.g., datamatrix in Solodovnikov & Schomann 2009; and work in progress) but only some of them are preliminarily thought to be synapomorphies: setose apical segments of the maxillary and labial palps; more or less distinctly elongate first antennomere; lack of conspicuous large macrosetae on mesoventrite; abdominal tergites III–VII with only one basal carina; and male sternite VIII without secondary sexual modifications, medially straight to very slightly concave. Within this tentative group, strongly dilated anterior tarsi and evenly setose head and pronotum are autapomorphies of Australotarsius .
Of the listed genera, only one species of Anchocerus ( Solodovnikov 2008) and five species of Acylophorus ( Newton & Thayer 2005) occur in Australia. But both species of Australotarsius can be easily distinguished from the Australian Anchocerus tenuipes ( Lea, 1929) externally by very broad anterior tarsi and setose head and pronotum. The same characters also easily separate Australotarsius from Acylophorus , and the latter genus, in addition, has a conspicuously narrow neck and a much more strongly elongate first antennomere.
According to the preliminary phylogeny of Staphylinini ( Solodovnikov & Schomann 2009), a quediinelike Australotarsius does not belong to “ Tanygnathinina sensu novo”, the lineage whose members superficially resemble “Quediina propria” and which is a predominant and the most abundant faunal element among Australian Staphylinini . Current Australian species of “ Quedius ”, and “ Heterothops ”, all members of “ Tanygnathinina sensu novo”, are not congeneric with the north temperate representatives of these genera; but some other, less species-rich Australian genera now formally in Quediina also belong to this group. From any species of “ Tanygnathinina sensu novo” Australotarsius differs in the structure of the mesoscutellum which has two (not one) transverse carinae, and in the shape of the paramere which is closely attached to the median lobe only at its base, and has distinct, strongly sclerotized and obtuse sensory peg setae (in “ Tanygnathinina sensu novo” the paramere is closely attached to the median lobe along the entire length of the latter, and lacks sensory peg setae, or, if those present, they are inconspicuous and sharp). By such combination of characters as very broad anterior tarsi, somewhat deflexed hypomera of the pronotum, evenly setose head and pronotum, strongly cuspidate teeth at the base of the mandibles, and well developed infraorbital ridges, Australotarsius can be easily distinguished from any other genus of Staphylinini recorded in Australia including the very poorly known and phylogenetically puzzling genera Lonia Strand, 1943 and Antimerus Fauvel, 1878 .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.