
Palangania sp.
|
publication ID |
https://doi.org/ 10.1206/577.1 |
|
persistent identifier |
https://treatment.plazi.org/id/A45FA45F-FFD7-E475-FED6-FD69E535FB6E |
|
treatment provided by |
Carolina |
|
scientific name |
Palangania sp. |
| status |
|
Figure 3J View Fig
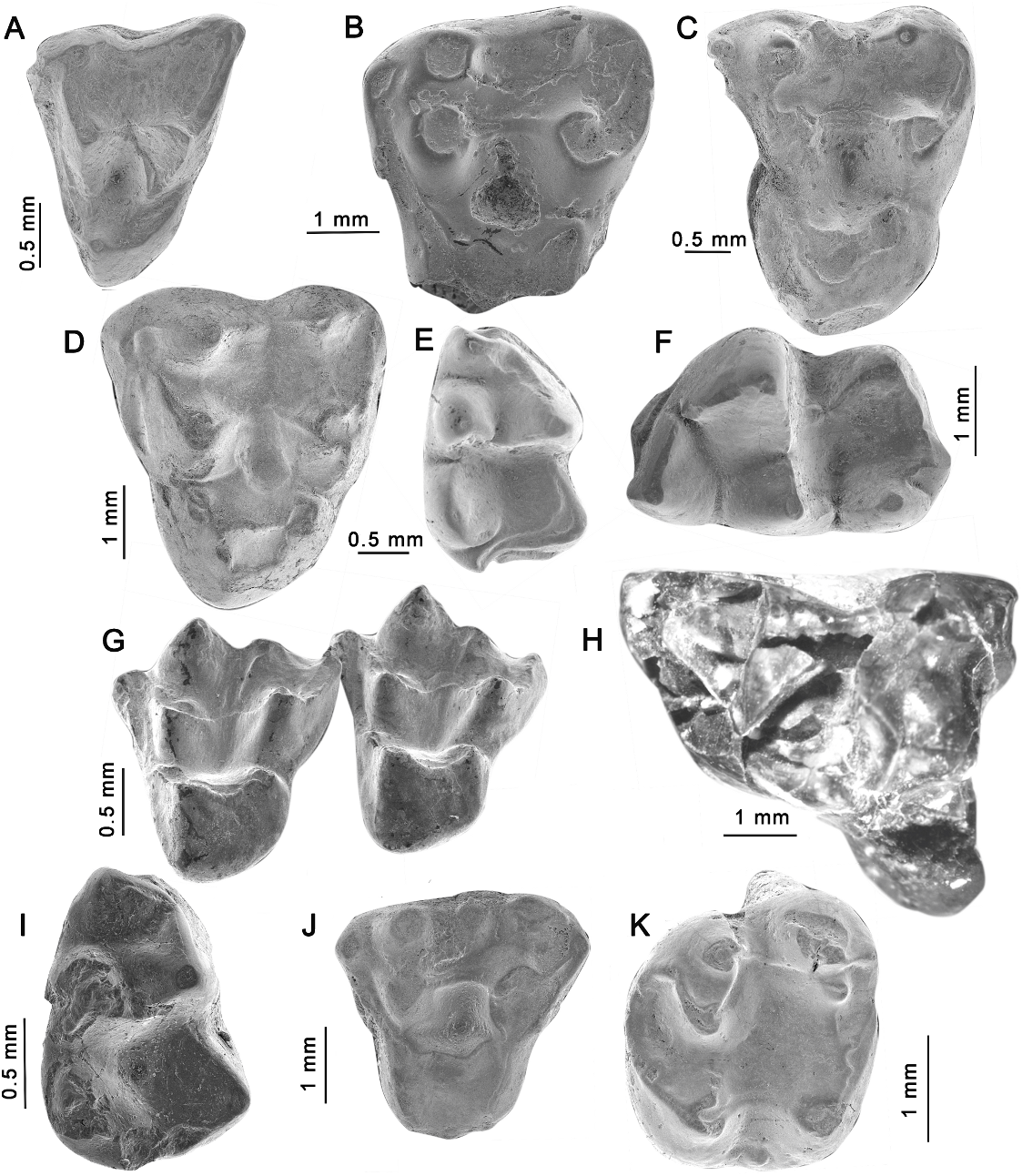
A single specimen (LIEB-PV 1106, left M?3; fig. 3J), an upper molar from La Barda, is complete but worn. The protocone is a prominent cusp and the trigon basin is deep and wide. The stylar shelf is reduced and StB and StD are placed close to the paracone and metacone, respectively; a lower StC is present. The centrocrista is almost linear, somewhat curved labially; the preparacrista ends on the anterolingual side of the StB and the postmetacrista is short. Due to the wear, it is not possible to confirm the presence of a metaconule, but a paraconule was already present. Also, the StC is relatively larger and the stylar shelf is more developed labiolingually. Although smaller and more generalized, this molar resembles the M3 of Palangania brandmayri .
Suborder Bonapartheriiformes Goin and Candela, 2004 Family Gashterniidae Marshall, 1987 Gashternia ctalehor Simpson, 1935 Figure 3K View Fig
The holotype of this species, AMNH 28533, a right mandibular fragment with m1–2 partially preserved is from the upper Paleocene of Patagonia. Several upper and lower molars attributed to this taxon were found at La Barda. Gashternia differs from other Polydolopimorphia in having lower molars with trigonid and talonid divided by a deep flexus anterior to the cristid obliqua. The metaconid is higher than the protoconid in m2–4 and becomes fused to the metaconid from m1 to m4. The anterobasal cingulum and hypoconulid are vestigial. Upper molars (e.g., LIEB-PV 1132, right M4; fig. 3K) have a deep lingual flexus separating the protocone from the hypertrophied and lingually displaced metaconule. StB and StD are the most developed stylar cusps while StC is lacking in all molars. The postmetaconular crest reaches the labial edge forming a wide sulcus.
The selenodont condition of Gashternia makes it one of the most singular South American marsupials. In the original description, Simpson (1948: 69) wrote:
this peculiar little jaw is quite unlike anything else known to me. … There is some suggestion that it may be a marsupial, chiefly the fact that the probabilities somewhat favor the presence of only three premolars, but this is not certain.
The material from Paso del Sapo led us to understand the morphology of all molars, including the uppers, and strongly supports the inclusion of Gashternia in the Metatheria.
Suborder Polydolopiformes Kinman, 1994 Family Polydolopidae Ameghino, 1897 Subfamily Polydolopinae Ameghino, 1897 Genus Polydolops Ameghino, 1897 Polydolops unicus , sp. nov.
HOLOTYPE: LIEB-PV 1252 , right maxillary fragment with P3–M3 (fig. 4a). P3 width 5 2.96; P3 length 5 2.46; M1 width 5 3.60; M1 length 5 2.89; M2 width 5 4.41; M2 length 5 2.6; M3 width 5 2.41; M3 length 5 2.24.
ETYMOLOGY: From the Latin unicus , ‘‘unique’’, recognizing the exceptionally derived morphology of its second upper molars.
REFERRED SPECIMENS: The holotype and LIEB-PV 1240 *, left maxillary fragment with M2–3; LIEB-PV 1241 *, left M1, LIEB-PV 1242 *, right M1, LIEB-PV 1243 *, right M1, LIEB-PV 1244 *, right, broken M1; LIEB-PV 1245 , left maxillary fragment with M1–2; LIEB-PV 1246 , left M1; LIEB-PV 1247 , left M2; LIEB-PV 1248 , left M2; LIEB-PV 1249 , left M2; LIEB-PV 1250 , left M2; LIEB-PV 1251 , right maxillary fragment with P3–M3; LIEB-PV 1253 , right M1; LIEB-PV 1254 , right M1; LIEB-PV 1255 , right M1; LIEB- PV 1256, right, broken M2; LIEB-PV 1257 , right M3; LIEB-PV 1258 , right M3; LIEB-PV 1212 *, left dentary with broken m1, complete m2, and roots of m3; LIEB-PV 1217 *, right dentary with m1–3; LIEB-PV 1259 *, left mandible with m1–2 and roots of p3; LIEB- PV 1260*, left mandible with m2–3; LIEB- PV 1261*, left m2; LIEB-PV 1262 *, right mandible with m1–2; LIEB-PV 1263 , left mandible with m1–2; LIEB-PV 1265 , left m2; LIEB-PV 1266 , left, broken m2; LIEB-PV 1267 , right mandible with broken p3, complete m1–2, and roots of m3; LIEB-PV 1268 , right mandible with m2; LIEB-PV 1269 , right mandible with m2; LIEB-PV 1270 , right mandible with m2; LIEB-PV 1271 , right mandible with broken m1, complete m2, and roots of m3; LIEB-PV 1272 , left m1; LIEB-PV
1273, right m1; LIEB-PV 1274, right m1; LIEB-PV 1275, right m2; LIEB-PV 1276, broken right m2; LIEB-PV 1277, right m3; LIEB-PV 1278, right m3.
GEOGRAPHIC AND STRATIGRAPHIC PROV- ENANCE: Specimens marked above with an asterisk (*) come from Laguna Fría, while the remainder are from La Barda; Tufolitas Laguna del Hunco Formation, early middle Eocene, western Patagonia.
DIAGNOSIS: Polydolops unicus (fig. 4) differs from other species of Polydolops in the following characters: large size, although smaller than P. mayoi and P. abanicoi ; shorter face, and higher and more robust maxillae and mandibles; M1 comparatively shorter; M2 broader than M1; the metacone and StB of M2 are fused and hypertrophied, thus forming a single cusp strongly projecting over the occlusal plane of the upper molars; m1–2 (and probably m3) relatively small and short in relation to the size of the mandibular corpus;
m1–2 hypoconid placed more anteriorly than in P. rothi .
DESCRIPTION: Abundant upper and lower teeth, mandibles, and maxillae of this new taxon are preserved, coming from both localities at Paso del Sapo. This new species is the most specialized polydolopine known to date; the functional significance of these adaptations are under study.
The holotype preserves most of the maxilla and cheek teeth. The maxilla is high in relation to other polydolopines, and it has a moderately developed infraorbital foramen located above the root of P3. The suture line between the maxilla and the anterior part of the jugal is preserved, showing that the jugal was vertically high. P3 is mesiodistally short and buccolingually wide, nearly as wide as the M1. The crown shows a large principal cusp and a smaller one anteriorly placed. The major cusp has two crests, anterior and posterior, and two ‘‘ridges’’, labial and lingual. The crests are not sharp compared to the ridges. The posterior root is broad, probably twice the size of the anterior root. A little-developed talon is present posterolingually. M1 is proportionally shorter and wider than P3, much wider in the posterior half. There are two cusp rows labially, not aligned but with an irregular pattern, especially in the more labial row; both rows join in a relatively large StB. A reduced paracone is seen on the lingual side of the StB. In the stylar shelf, the internal row is formed by the StC and StD aligned and subequal in size and height, and the StE somewhat lower than the StB. A metacone is observed between StD and StE, with the base fused to these stylar cusps, and the metacone is almost the height of StE. The labial row is formed by four accessory cuspules. M1 exhibits two lingual lobes, with a paraconule in the anterior one. Three subequal cusps are observed in the anterior half of the posterior lobe: the first is probably the protocone, followed by an accessory cusp, and the third is probably the metaconule, but the homologies are difficult to assess.
M2 is an odd tooth, very derived and unique among the Metatheria. Its degree of specialization is even more surprising considering that M1 and M3 do not show the same specialization. The unusual buccolingual width is clearly greater than in M1, and more than twice that of M3. The lingual edge of M2 is aligned with the lingual edges of M1 and M3, thus leaving the labial edge projecting beyond the labial edge of M1. Another surprising character is the possible presence of four roots, with a large lingual and three labial roots. It is possible that the anterolabial and centrolabial roots are parts of the same root with two lobes. But the most remarkable trait is the enormous cusp placed centrolabially; this hyperthrophied tooth has modified the total volume of the tooth as well as the supporting root; as a result, the labial roots have developed an additional lobe. This huge cusp appears to be formed by the fusion of the metacone and StD. The lingual margin of M2 is relatively linear with poorly developed paraconule and metaconule that have metaconular crests. The trigon basin is rugged. The M3 is reduced and simplified compared to M1–2, being generally subtriangular in shape, longer lingually than labially.
There is remarkable variability in the size and robustness of the mandibles, probably suggesting sexual dimorphism. The anterior part of the mandibles is not preserved in any specimen of P. unicus , thus there is no information on the incisors and anterior premolars. The p3 is only known in two specimens but only the base was preserved; it was apparently short and robust. The lower molars are compact, robust, and slightly hypsodont labially (labial side clearly higher than the lingual side). The m1–2 are subequal in length and width, while m3 is longer and narrower. The occlusal surface was crenulat- ed. The m1 shows a very reduced and short trigonid, with two low cusps. Behind the trigonid, there are three lingual and three labial cusps. The labial cusps are shallow and robust, the posterior (hypoconid) being the most robust. The m2 is shorter and shallower than the m1, with just four lingual and four labial cusps, among which the metaconid (as in other polydolopines) is the highest and more robust. The most striking trait in m2 is the anterior position of the hypoconid; this trait was incipient in Polydolops rothi , but has a high degree of specialization in P. unicus . This trait clearly distinguishes P. unicus from any other polydolopine species. In polydolopines, the posthypocristid is almost parallel to the posterior margin of the tooth, slightly curving forward to meet the hypoconid. To the contrary, in P. unicus (and in a much lesser degree in P. rothi ), the posthypocristid is oblique to the dental axis, slightly concave. This odd morphology is certainly related to the occlusion with the huge cusp of M2, and due to the hypertrophy of this cusp the interdental space between m2–3 leave a place for the occlusal closure.
REMARKS: Polydolops unicus is the most specialized polydolopine known. This is especially noteworthy in light of the specialization of the entire Polydolopinae , whose morphology has diverged considerably from the tribosphenic pattern. On the other hand, P. unicus was previously unknown from other stratigraphic levels in Patagonia, and its relative abundance in the Paso del Sapo mammal assemblages is surprising, thus characterizing the faunal associations. Therefore, P. unicus may be considered as a fossil guide (see below). The only known polydolopine that may be treated as a potential ancestor of the morphology of P. unicus is P. rothi , which shows a derived pattern in the molar morphology having: (1) M2 with enlarged StD, lingually inclined and coalescent with the metacone forming a transversal axis to the dentary; (2) m2 with the hypoconid anteriorly displaced with consequent development of the slightly concave hypocristid. These characters may allow P. unicus and P. rothi to be considered different genera, but this hypothesis should be tested in an integrated analysis of the whole family.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
Palangania sp.
| Tejedor, Marcelo F., Goin, Francisco J., Gelfo, Javier N., López, Guillermo, Bond, Mariano, Carlini, Alfredo A., Scillato-Yané, Gustavo J., Woodburne, Michael O., Chornogubsky, Laura, Aragón, Eugenio, Reguero, Marcelo A., Czaplewski, Nicholas J., Vincon, Sergio, Martin, Gabriel M. & Ciancio, Martín R. 2009 |
Polydolops unicus
| Tejedor & Goin & Gelfo & López & Bond & Carlini & Scillato-Yané & Woodburne & Chornogubsky & Aragón & Reguero & Czaplewski & Vincon & Martin & Ciancio 2009 |
Polydolopiformes
| Kinman 1994 |
Polydolopidae
| Ameghino 1897 |
Polydolopinae
| Ameghino 1897 |
Polydolops
| Ameghino 1897 |