
Cricotopus varicornis Drayson & Cranston
|
publication ID |
https://doi.org/10.11646/zootaxa.3919.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:218630EE-6BF7-4E35-A8F6-9E8260D60FA0 |
|
DOI |
https://doi.org/10.5281/zenodo.6122653 |
|
persistent identifier |
https://treatment.plazi.org/id/E290B6D9-D10A-447A-9832-AFAB8948DC45 |
|
taxon LSID |
lsid:zoobank.org:act:E290B6D9-D10A-447A-9832-AFAB8948DC45 |
|
treatment provided by |
Plazi |
|
scientific name |
Cricotopus varicornis Drayson & Cranston |
| status |
sp. nov. |
Cricotopus varicornis Drayson & Cranston View in CoL sp. n.
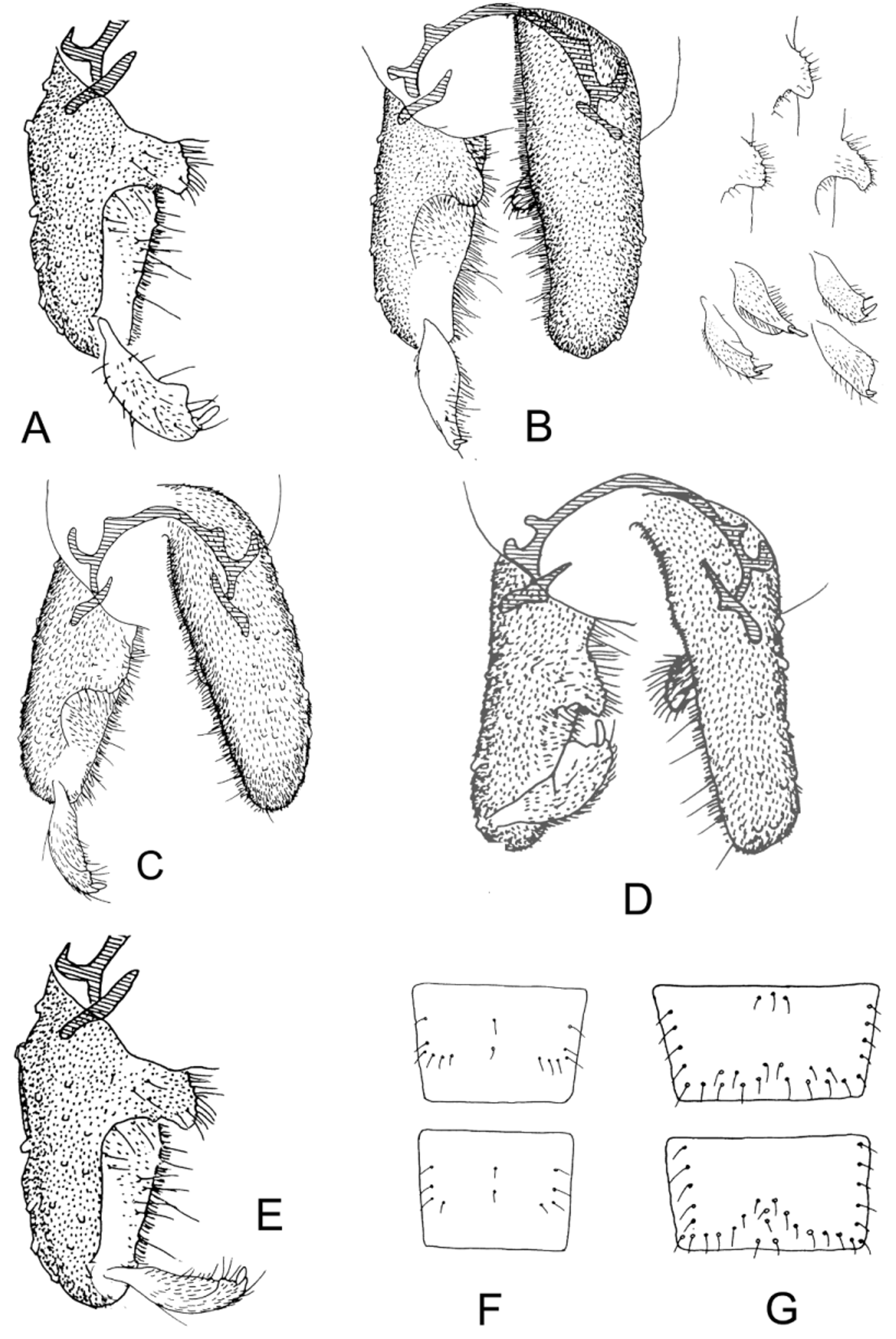
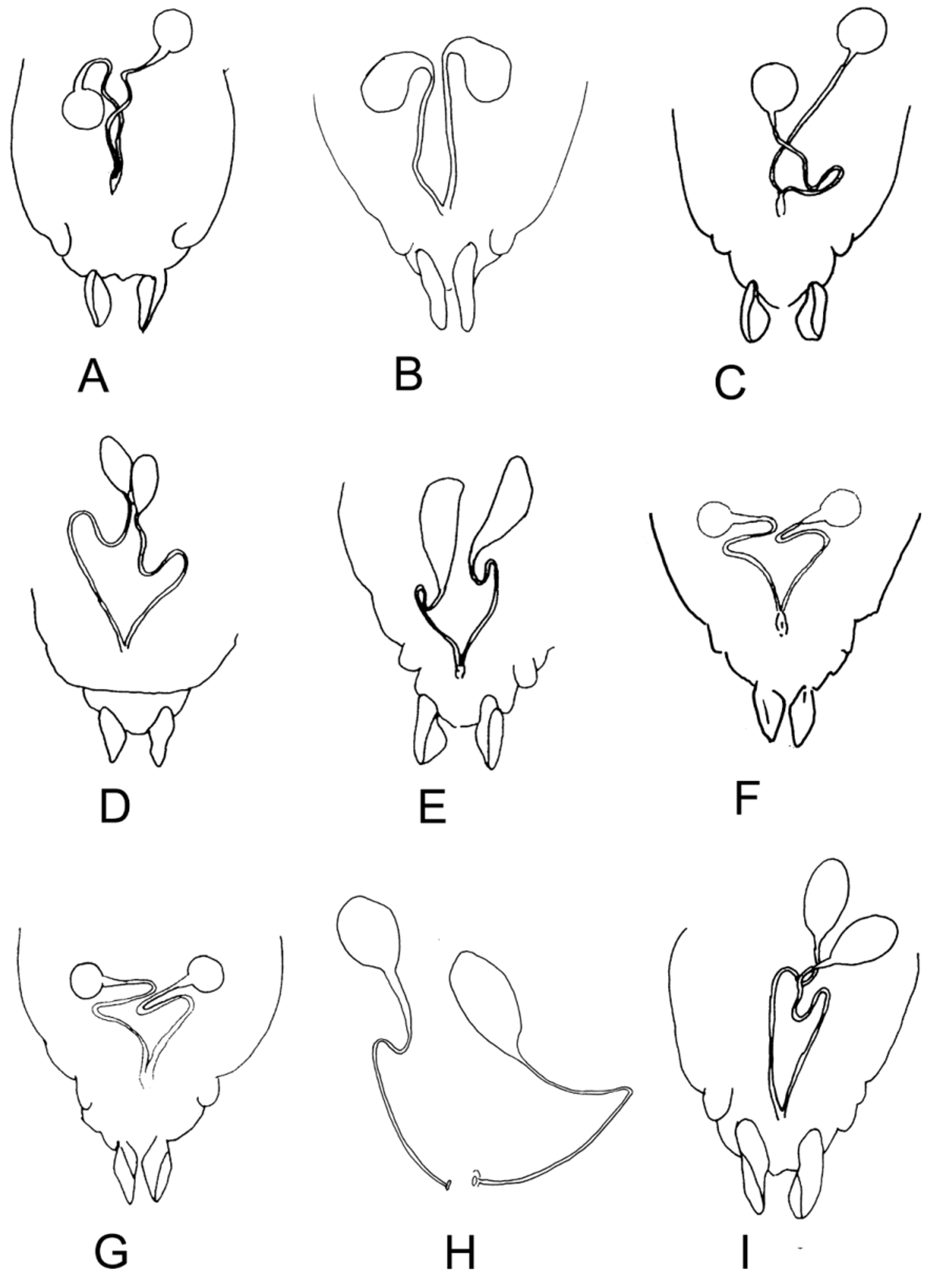
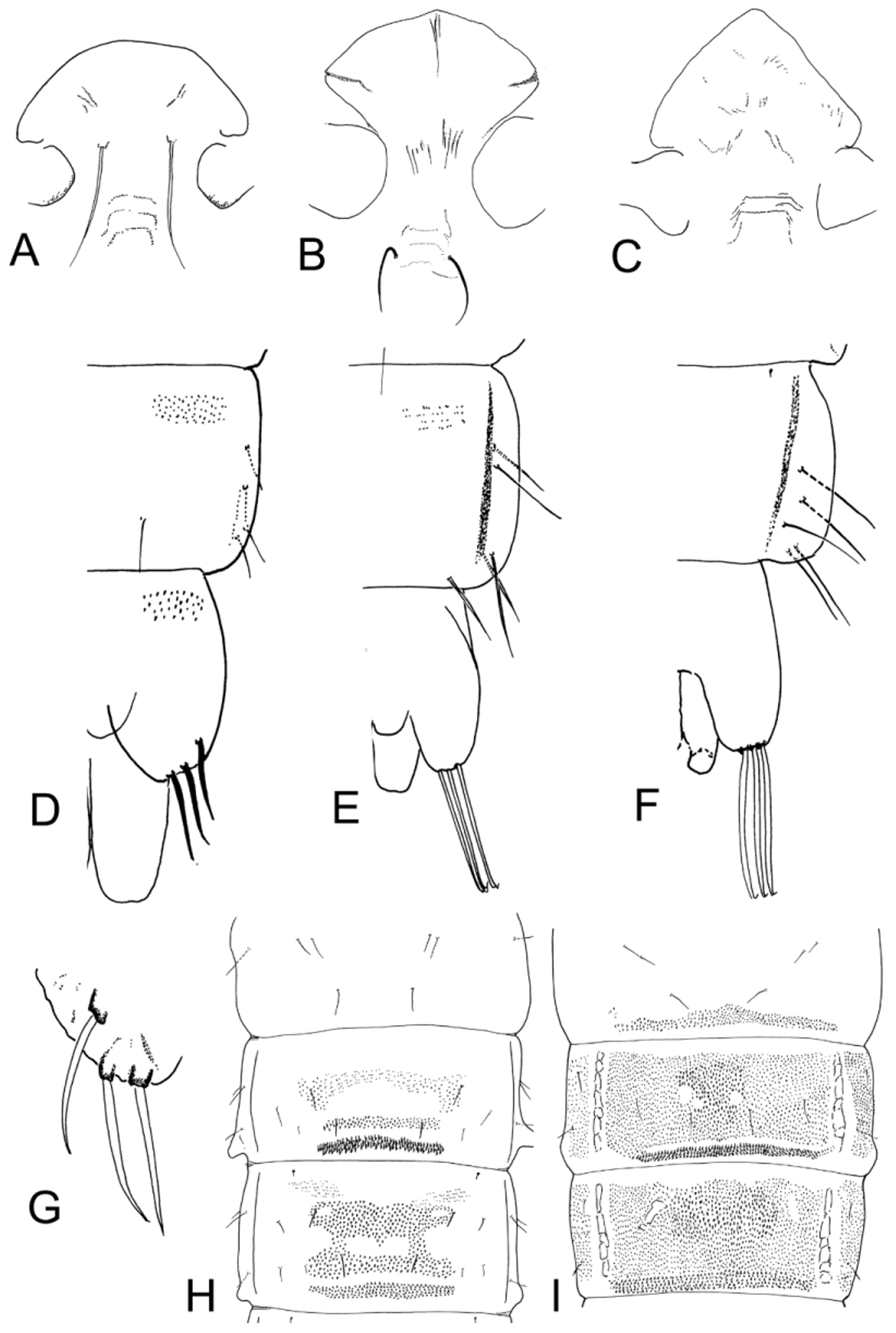
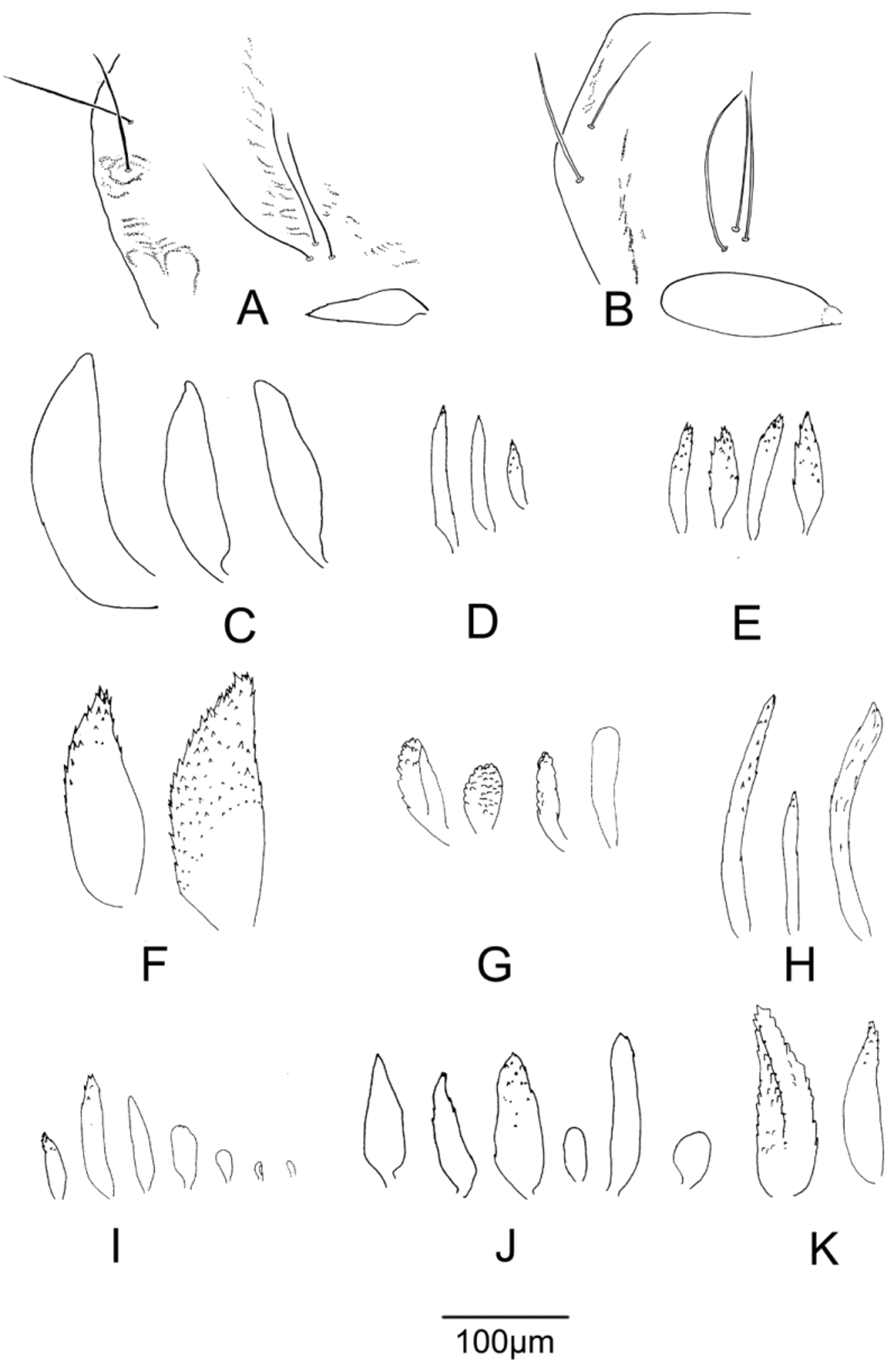
( Figs. 1 View FIGURE 1 I, 3D,F, 5I, 7A,J, 9D, 12B)
urn:lsid:zoobank.org: act :E290B6D9-D10A-447A-9832-AFAB8948DC45
Cricotopus View in CoL ‘sp. D’ Drayson, 1992: 102 (nec larva, misassociated)
Cricotopus View in CoL ‘sp. V’ Drayson, 1992: 73
Cricotopus View in CoL “ varicornis ” Cranston sp. nov. in Cranston, 1996: 87 [Invalid; author states ‘not formal publication for nomenclatural purposes]
Cricotopus View in CoL “cooki” sp. nov. Drayson & Cranston, in Cranston, 1996: 86 [Invalid; author states ‘not formal publication for nomenclatural purposes’]
Type material. Holotype: P♂, AUSTRALIA; ACT, Corin Rd , Gibraltar Falls, 35°28'S 148°55'E, 23.ix.1991 (Drayson). Paratypes: as holotype, except ♂, 2 Pe/♂, 2♀, 2 Pe/♀, 27 Pe, 5 L, 2 Le/P.
Additional material examined 1. Assigned to form varicornis .: ♂, Le/P♂, Le/P♀, 2P♀, 3LP, 7Pe,, as type except 13.x.1993 (Cranston); ♀, as type, except 20.x.1993 (Cranston); L as type except 23.x.1993. 2. Assigned to form cooki: Vic: Pe, Wodonga, House Ck., upstream, 36°10'S 146°52'E, 19.ii.1989 (Cook); 1Pe, Wodonga, Middle Ck., Street's Rd, 36°11'S 146°56'E, 26.ii.1990 (Cook); 11 Pe, Wodonga, Middle Ck., Boyes Rd., 36°13'S 146°54'E, 7.xii.1989 (Cook); 3Pe, Wodonga, Middle Ck., Beechworth Rd, 36°15'S 146°50'E, 5.i.1990 (Cook).
Molecular material. 2L, P♀, ACT , Paddy’s R., 35°27'S 149°01'E, 22.iv.2013 (Cranston) (Mv-ACTPR6-8 - cooki); NSW: P♀ Pipers Ck., 36°23'S 148°26'E, 2.xii.2010 (Cranston) (Mv-KNPCric7- varicornis ).
Description. MALE ( Fig. 1 View FIGURE 1 I) (n = 2 + 3 pharate). 2.7–3.3 mm.
Head. Ant 680–744 µm; Fl 1–12, 350–400 µm, Fl 13, 275–400 µm; A.R. 0.7–0.96. Palp 308–324 µm. Fr 2, Po 3. Clyp moderately setose.
Thorax. Background yellow-brown, with mid-brown scutal vittae and postnotum distinctive. Laps 4–7, Ac 9–17, Dc 12–25, Pa 5–6, Scts 7–12.
Wing. 1.8 mm. Sq 5–9, R 0–5.
Legs. Unmeasurable; fore tibia pale from basal area to mid-tibia, otherwise weakly to distinctly dark.
Abdomen. T I and IV very pale; broad anterior pale band on TII, otherwise segments mid-to dark brown. TIII and VI with lateral setal bands not connected ( Fig. 3 View FIGURE 3 G).
Hypopygium ( Fig. 3 View FIGURE 3 D). Gcx 170–200 µm, iv directed postero-medially, very setose, without hyaline apex. Gst 70–88 µm, about 1/2 (0.4–0.5) Gcx; crista dorsalis high, elongate.
FEMALE (n=4, pharate). As in male except: 3.3–3.6 mm.
Head. Ant 301–316 µm. Palp 382–394 µm.
Thorax. Laps 5–7, Ac 12–23, Dc 12–22, Pa 4–6, Scts 8–10.
Wing. 1.9–2.0 mm. Sq 2–5, R + R 1 10–19; R4+ 5 8–11.
Legs. Brown with paler section from near base to middle of fore-tibia, indicated also on mid- and hind legs.
Genitalia. Spermathecae comprising brown, ovoid capsules with narrow "neck" and recurved ducts ( Fig. 4 View FIGURE 4 I).
PUPA. 2.9–5.4 mm, pale brown to almost hyaline.
Cephalothorax. Moderately rugose dorsally. Th 26–150 µm; hyaline to brown pigmented, very variable in shape, with or without apical scales ( Fig 7 View FIGURE 7 A,J). Fs 26–70 µm, on frons.
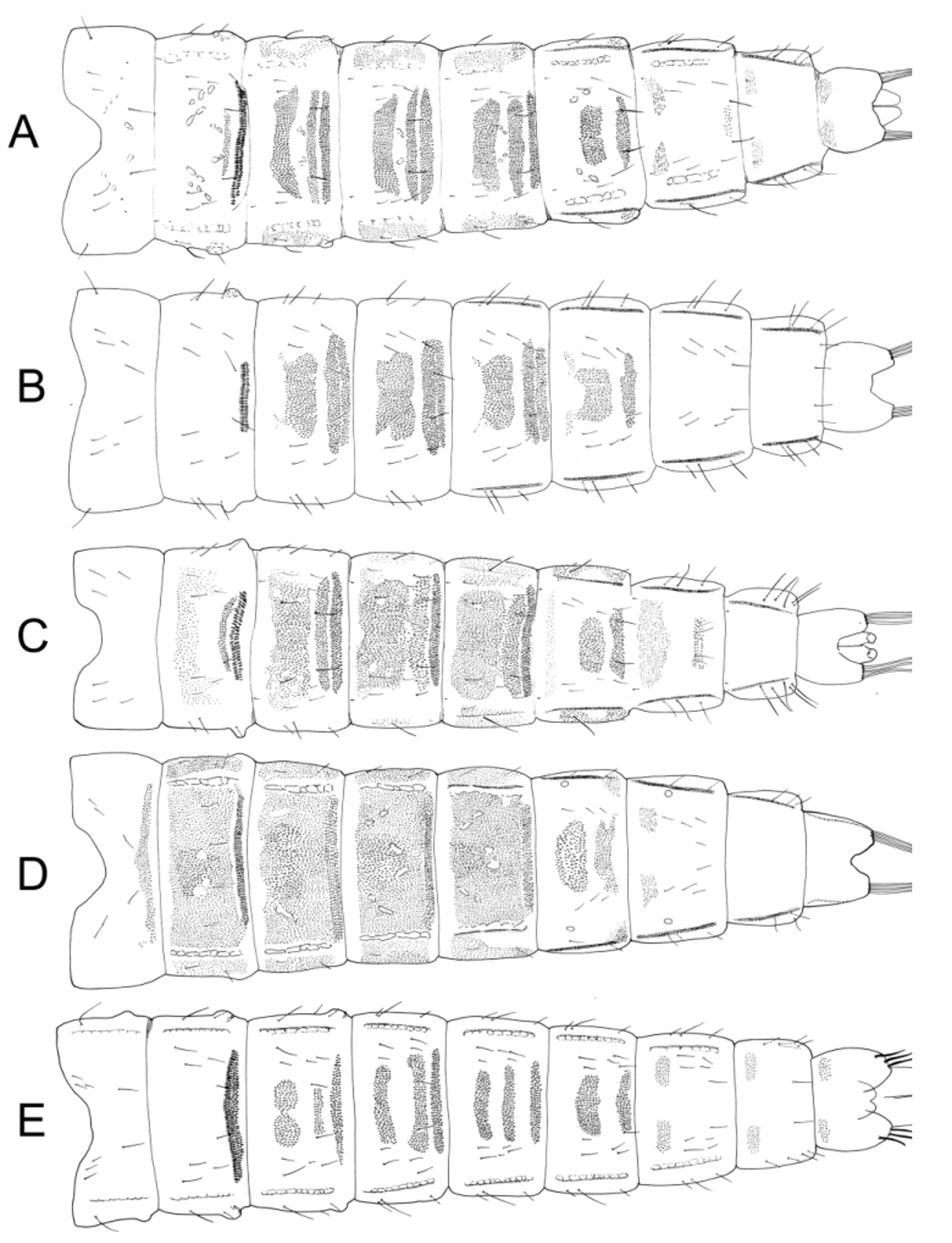
Abdomen ( Fig. 9 View FIGURE 9 D). PSB on II, smaller on III. Hook row about 1/2 of segment (0.42–0.56). Spinule fields present on TI and anterior to hook row on TII ( Fig. 5 View FIGURE 5 I). Paratergite spinulose on II–V. Ls 3 VIII short, 56–80 µm, <1/5 segment width (0.12–0.20).
Ms 110–220 µm, less than 1/15 length of abdomen (0.05–0.06). Extensive spinulation on sternites II and III.
4TH INSTAR LARVA (Fig. 12B, C) (n=3) Length not measurable. H.l. 470–480 µm very pale with pigmented areas including posterior margin dark brown; abdomen yellow with strong blue colour; procercus hyaline, with mid-brown marking. Anterior parapod claws pale yellow; posterior parapod claws darker yellow.
Head. Ant 74–78 µm, 1st 37–40 µm, 2–5 32 –37 µm; A.R. 1.0–1.2. Md 130–140 µm with smooth inner and strongly or weakly crenulate outer margin, dark brown to notch and distal mola, posterior ½ to ⅔ pale yellow. Mentum 107–117 µm, middle tooth wide, perhaps complex, 22–25% mentum width, pale brown posteriorly, midto dark brown anteriorly; with 6 pairs lateral teeth on even slope.
Abdomen. Pc 20 µm, A.s. 500 µm.
Remarks. The variation in morphology of this taxon, reflected in the epithet ‘ varicornis’, has caused confusion since the earliest material was collected in the late 1980s. Two near identical pupal exuvial types exist, one with a variably shaped but always short (less than 100 µm) hyaline thoracic horn and always smooth or nearly so, and another with thoracic horn longer (to 150 µm), darker pigmented, and apically rugose. The former was named informally as ‘ varicornis’, the latter as ‘ cooki’. Molecular data associate larvae with a pupal morphotype that are distinctive by their wide, seemingly complex, median mental tooth, antennal segment 2 near wedgeshaped, shorter than the 3rd segment, and antennal ratio closer to 1.0. However, Drayson’s illustrated purported larva of ‘ varicornis’ (1992: 102) differs with narrow median mental tooth, antenna with segment 2 cylindrical and longer than 3rd and A.R. greater than 1.5. However even recognising this error, significant variation exists also amongst larvae otherwise with ‘diagnostic’ morphology, not least in features that have been considered central to larval recognition. Thus the outer mandibular crenulation ranges from distinct (‘ varicornis’) to slight or absent (‘ cooki’), and the extent of mandibular pigment also varies. Adult males associated with each pupal type, though few, appear identical including in fine details of the hypopygium.
Based on the material currently available, we consider all larvae and pupae to vary in characters considered otherwise to be diagnostic. Evidence for conspecificity derives from other morphological features. Molecular evidence, although based on few specimens, shows ‘ cooki’ is sister to a solitary pupa identified as varicornis (Krosch et al., submitted). Should additional material and molecular data clarify that two species are involved, cooki is available for specimens that conform to the larval and pupal features described above. For this reason we restrict the type-series to the morphotype pupa with shorter hyaline thoracic horn, but with length and pigment variation not extending to the ‘ cooki’ form, and a larval morphotype with crenulate outer mandible and with extended pigment.
We treat each as a form, and include both in the larval and pupal keys.
C. varicornis inhabits clean, cool, south-eastern Australian streams.

| NSW |
Royal Botanic Gardens, National Herbarium of New South Wales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cricotopus varicornis Drayson & Cranston
| Drayson, Nick, Cranston, Peter S. & Krosch, Matt N. 2015 |
Cricotopus
| Cranston 1996: 87 |
Cricotopus
| Cranston 1996: 86 |
Cricotopus
| Drayson 1992: 102 |
Cricotopus
| Drayson 1992: 73 |