
Albunea groeningi, BOYKO, 2002
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2002)272<0001:AWROTR>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/A23087F4-FED1-FE91-F2B6-C46271467EF7 |
|
treatment provided by |
Felipe |
|
scientific name |
Albunea groeningi |
| status |
sp. nov. |
Albunea groeningi View in CoL , new species Figures 96 View Fig , 97 View Fig
Albunea symnista [sic]: Gordon, 1938: 187 (part)*. – Miyake, 1978: 152–154, figs. 59, 60a* (not Albunea symmysta ( Linnaeus, 1758)) View in CoL .
Albunea symmysta: Miyake et al., 1962: 125 View in CoL (part; Toyama Bay material) (not Albunea symmysta ( Linnaeus, 1758)) View in CoL .
Albunea steinitzi: Serène and Umali, 1965: 97– 102 View in CoL , pl. 1, fig. 2, pl. 2, fig. 2, pl. 3, figs. 3–4b, pl. 4, fig. 2, textfigs. 1b, 2b, 4b, 5c, 6c, c̍, 7a, 9b, c*. – Haig, 1974: 447 (list) (not Albunea steinitzi Holthuis, 1958 View in CoL ).
? Albunea symnista [sic]: Kikuchi, 1932: 10 (? not Albunea symmysta ( Linnaeus, 1758)) View in CoL .
MATERIAL EXAMINED: Japan: MikawaIsshiki, Aichi Prefecture, Honshu , Sept. 1, 1941, coll. T. Sakai: 1 Ƌ, 9.4 mm cl, holotype ( RMNH ex 32052), 1 ♀, 12.5 mm cl, allotype ( RMNH 32052 About RMNH ) ; Sakai , Tottori Prefecture, March 1, 1964, coll. T. Senda: 1 ♀, 16.4 mm cl ( ZLKU 12836 ) ; Fushiki, Toyama Bay , Aug. 6–26, 1950, coll. Hori and Mori: 1 Ƌ, 8.7 mm cl, 1 ♀, 7.0 mm cl, paratypes ( ZLKU 5125–5126 ) ; Mimase, Kochi City, Tosa Bay , Shikoku Island , Dec. 24, 1959, coll. K. Sakai: 1 ♀, 17.0 mm cl, paratype ( ZLKU 7438 ) ; Tosa Bay, Shikoku Island , Feb. 19, 1960, coll. S. Nakayama: 1 Ƌ, 11.6 mm cl, paratype ( ZLKU 7511 ) .
Taiwan: ‘‘ Formosa ,’’ coll. M. Maki: 1 ♀, 13.8 mm cl, paratype ( USNM 59074 About USNM ) ; Kaohsiung, 25 m, June 7, 1992, coll. unknown: 2 Ƌ, 9.2–9.8 mm cl, 1 ♀, 11.2 mm cl, paratypes ( NTOU) ; ‘‘ Southern Taiwan ,’’ 30 m, Jan. 1, 1992, coll. unknown: 1 Ƌ, 8.8 mm cl, 1 ♀, 9.2 mm cl, paratypes ( NTOU) ; Tung Shiao, Miao – Li County, Aug. 30, 1999, coll. unknown: 1 ♀, 8.4 mm cl ( NTOU) ; Tan Shui, Taipei County , June 20, 1999, coll. unknown: 1 Ƌ, 4.1 mm cl ( NTOU) .
Philippines: Busuanga , Palawan, May 10–30, 1963, coll. J. E. Norton and F. E. Dayrit: 1 Ƌ, 6.4 mm cl, paratype ( NMCR 1151 b) ; Calapan, Mindoro , Dec. 1932 – Jan. 1933, coll. P. de Mesa: 1 ♀, 7.9 mm cl, paratype ( MCZ 9625 About MCZ ) .
Singapore: Singapore , coll. unknown: 1 Ƌ, 9.6 mm cl, paratype ( BMNH 1937.6.1.8) ; Morib Beach, Selengar, March 8, 1993, coll. A. Sasekumar: 1 oviger, 10.7 mm cl, paratype ( ZRC 1995.571 View Materials ) .
Malaysia: Bedok, Feb. 7, 1959, coll. P. Yeoh: 1 Ƌ, 10.8 mm cl ( ZRC 2000.1799).
Vietnam: Sta. 51, off Nhatrang, 15 m, Jan. 14, 1960, coll. R/V ‘‘Gallardo’’: 1 Ƌ, 3.8 mm cl ( ZMUC 2717 View Materials ) ; Sta. 215, off Nhatrang, 16 m, March 7, 1960, coll. R/V ‘‘Gallardo’’: 1 Ƌ, 3.0 mm cl ( ZMUC 2718 View Materials ) ; Sta. 54, off Nhatrang, 14 m, Jan. 14, 1960, coll. R/V ‘‘Gallardo’’: 1 ♀, 3.7 mm cl ( ZMUC 2719 View Materials ) ; Sta. 220, off Nhatrang, 16 m, March 7, 1960, coll. R/V ‘‘Gallardo’’: 1 unsexable, unmeasurable specimen ( ZMUC 2721 View Materials ) .
Australia: Western Australia: ‘‘ Northwest Australia,’’ coll. unknown: 1 Ƌ, 6.0 mm cl, paratype ( BMNH 1932.11.30.65) ; Rosemary Island, Dampier Archipelago , 7 fms (= 12.8 m), Aug. 22, 1963, coll. F.R.V. Lancelin: 3 Ƌ, 9.2–11.4 mm cl, 2 ♀, 10.6–11.3 mm cl, paratypes ( WAM 23386) ; Bernier Island, Shark Bay , May 16, 1960, coll. R. W. George: 1 ♀, 13.4 mm cl, paratype ( WAM 23391) ; Shark Bay , Feb. 1963, coll. L. R. Thomas: 1 Ƌ, 12.9 mm cl, paratype ( WAM 23395) ; North Island , 17–25 fms (= 31.1– 45.7 m), May 1965, coll. R. Seabrook: 1 Ƌ, 12.9 mm cl, paratype ( WAM 23396) ; Mission Bay, Napier Broome Bay , 1–3 m, Aug. 14, 1991, coll. L. Wharsh: 2 Ƌ, 6.4–8.3 mm cl, 1 ♀, 8.1 mm cl, paratype ( WAM 20667) ;
Queensland: Ball Bay, May 24, 1964, coll. Zoology Department of the University of Queensland: 2 Ƌ, 10.5–12.4 mm cl, 4 ♀, 7.8–15.4 mm cl, paratypes ( WAM 23389) ; Rockhampton, Keppel Bay, 23°10̍S, 150°55̍E, 9 m, Sept. 6, 1967, coll. B. J. Smith: 1 unsexable, unmeasurable specimen ( MOV J47044 View Materials ) ; Lucinda, Hinchinbrook Channel , 18°29̍S, 146°16̍E, Oct. 1969, coll. E. Gossner: 1 Ƌ, 11.0 mm cl, paratype ( MOV J14552 View Materials ) ; Rudder Reef, 30 mi northeast of Mossman , 16°11̍S, 145°40̍E, Oct. 1973, coll. R. J. Plant: 1 Ƌ, 8.5 mm cl, para type ( MOV J44728 View Materials ) ; Rockhampton, Keppel Bay, 23°10̍S, 150°55̍E, 9 m, Sept. 6, 1967, coll. B. J. Smith: 1 Ƌ, 14.1 mm cl, paratype ( MOV J44732 View Materials ) ; Dunk Island , 17°57̍S, 146°09̍E, coll. E. J. Banfield: 2 Ƌ, 8.9–9.2 mm cl, paratypes (AM P5340) ; off North Head, Port Denison , 20°01̍S, 148°15̍E, coll. E. H. Rainford: 1 Ƌ, 14.4 mm cl, paratype (AM P7029) ; Queen’s Beach, Bowen , coll. E. N. Rainworth: 1 Ƌ, 9.1 mm cl, paratype ( QM W143 ) ; Kinkuna National Park, 20 km south of Bundaberg , 25°00̍S, 152°30̍E, April 1992, coll. M. Hines: 1 unsexable specimen, 12.5 mm cl ( QM W18006) ; Weipa , July 1961, coll. G. Webster: 1 Ƌ, 9.1 mm cl, 1 ♀, 9.6 mm cl, paratypes ( QM W2221 ) ; Bowen , Feb. 27, 1934, coll. unknown: 1 Ƌ, 11.6 mm cl, paratype ( QM W489 ) ; Townsville , Oct. 7, 1938, coll. G. Coates: 1 ♀, 10.5 mm cl, paratype ( QM W873 ) ; Victoria: Mud Island, Port Phillip Bay, April 1977, coll. R. Willington: 1 ♀, 13.9 mm cl, paratype ( MOV J40194 View Materials ) .
DIAGNOSIS: Carapace wider than long, covered with lightly setose grooves. Anterior margin with 8–11 spines on either side of ocular sinus. Setal field with narrow lateral elements and concave anterior margin. CG1 with separate posterior lateral elements; CG4 with two to six short, oblique medial elements (rarely absent) between longer supralateral elements of CG4; CG5 present as two short transverse elements; CG6 and CG7 separate; CG8 broken; CG11 absent. Rostrum present, reaching posterior margin of ocular plate. Ocular plate triangular. Distal peduncular segments dorsoventrally flattened and subtriangular in shape, tapering at tip, approximated at base of mesial margins, lateral margins convex except slightly concave at tip, mesial margins straight. Cornea at tip. Dactylus of pereopod II with heel produced and acute. Dactylus of pereopod III with heel thin, produced and acute. Dactylus of pereopod IV with produced, subquadrate heel. Telson of male elongate, ovate, tip rounded with medial indentation, dorsal surface inflated medially, with medial row of long thickened setae; lateral margins decalcified. Telson of female similar to male, evenly calcified.
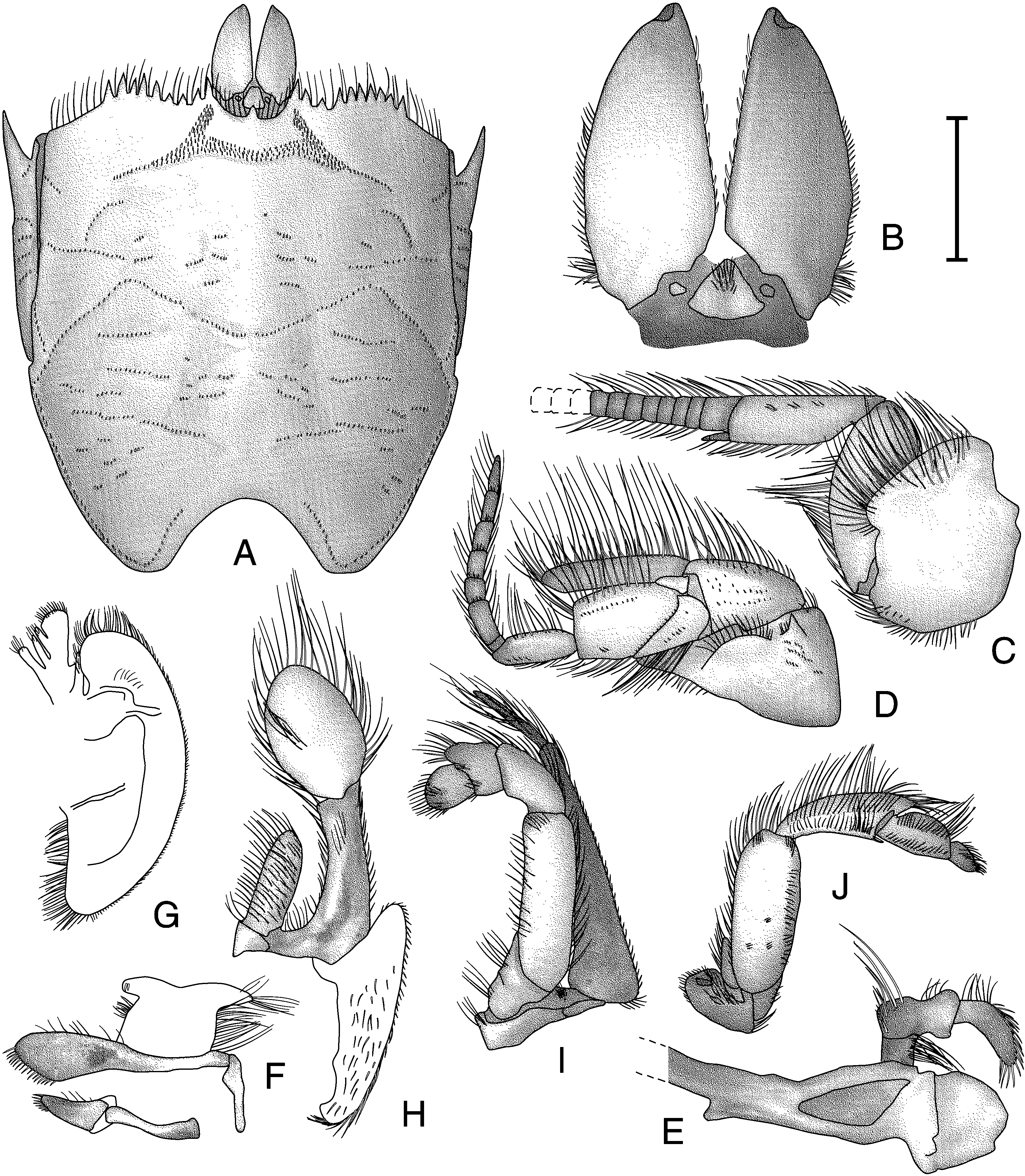
DESCRIPTION: Carapace (fig. 96A) wider than long. Anterior margin concave on either side of ocular sinus, becoming convex lat erally with 8–11 large spines (n = 6) on each side along length. Rostrum as small acute tooth, reaching proximal margin of ocular plate. Ocular sinus smoothly concave. Frontal region smooth; setal field narrow anteriorly and posteriorly; posterior lateral elements reduced to narrow bands of setae. CG1 parallel to anterior margin of carapace, convex, sinuous, strongly crenulate, divided into medial fragment and curved, posteriorly displaced lateral elements. Mesogastric region smooth; CG2 present as none to two short elements; CG3 broken into four widely separated short elements between posterior lateral elements of CG1; CG4 with two to six short, oblique medial elements (rarely absent) between longer supralateral elements of CG4. Hepatic region smooth, with oblique setose groove at median of lateral margin. Epibranchial region generally triangular, smooth; posterolateral margin with single oblique row of short setae. Metagastric region smooth; CG5 present as two short transverse elements. CG6 strongly crenulate, strongly anteriorly concave medially and sloping out to anteriorly convex lateral thirds. CG7 almost transverse, not reaching lateral margins of median segment of CG6. Cardiac region smooth; CG8 present as two to four short medial elements and two longer lateral elements. CG9 present as two to four short lateral grooves with broad gap at midline. CG10 present as two long lateral elements. CG11 absent. PostCG11 element absent. Branchial region with three to six short, oblique rows of setae in anterior half. Posterior margin deeply and evenly convex, with submarginal groove reaching twothirds up margin of posterior concavity. Branchiostegite with short anterior submarginal spine; anterior region with scattered, short, transverse lines ventral to linea anomurica; with many short rows of setae and sparsely covered with long plumose setae ventrally; posterior region membranous, with numerous irregular fragments and sparsely covered with long plumose setae.
Ocular plate (fig. 96B) triangular, with shallow median indentation; median peduncular segments present as small ovate calcified areas lateral to ocular plate. Distal peduncular segments elongate, subtriangular, with convex lateral and straight mesial mar gins, cornea covering lateral portion of distal tip; lateral margin with faint notch onefourth distal from base; mesial margins approximated at base; mesial margins with long plumose setae; tuft of plumose setae at proximolateral ventral angles and ventromedial oblique row of plumose setae extending from tuft to threefourths length of segment.
Antennule (fig. 96C) with segment III narrow proximally, expanding distally to three times proximal width; plumose setae on dorsal and ventral margins and sparsely scattered on lateral surface; dorsal exopodal flagellum with 104–127 articles (n = 6), long plumose setae on dorsal and ventral margins; ventral endopodal flagellum with two articles (n = 6), plumose setae on dorsal and ventral margins. Segment II medially inflated in dorsal view, with plumose setae on dorsal and ventral margins. Segment I wider than long, unarmed; dorsal third of lateral surface rugose with long plumose setae; long plumose setae on dorsal and ventral margins.
Antenna (fig. 96D) with segment V approximately two times longer than wide, with long plumose setae on dorsal and ventral margins and scattered on lateral surface; flagellum with seven articles (n = 6), long plumose setae on dorsal, ventral, and distal margins. Segment IV expanded distally, with long plumose setae on dorsal, ventral, and distal margins, and row of short setae on dorsolateral surface. Segment III with long plumose setae on ventral margin; short simple setae on dorsal margin and in short row on surface. Segment II short, widening distally, rugose, with plumose setae on margins and short simple setae scattered on lateral surface; antennal acicle long, thin, and exceeding distal margin of segment IV by onethird length of segment IV, with long plumose setae on dorsal margin. Segment I rounded proximally, flattened ventrolaterally, with long plumose setae on dorsal and distoventral margins, and short simple setae in short row on surface rugae behind spine; lateral surface with acute spine dorsodistally; low semicircular dorsolateral lobe ventrodistal to spine, margin of lobe with long plumose setae; segment with ventromesial antennal gland pore.
Mandible (fig. 96E) incisor process with three teeth; cutting edge with one tooth. Palp threesegmented, with plumose setae on margins and long, thick, simple setae arising from bend in second segment and on distal margin of terminal segment.
Maxillule (fig. 96F) distal endite proximally narrow, widening to inflated distal end, with thick simple setae on distal margin and thin simple setae on dorsal margin. Proximal endite with thick simple setae on distal margin. Endopodal external lobe truncate distally and curled under; internal lobe reduced with two thick setae at distolateral margin.
Maxilla (fig. 96G) exopod evenly rounded, with plumose setae along distal margin. Scaphognathite bluntly angled on posterior lobe, with plumose setae.
Maxilliped I (fig. 96H) epipod with plumose setae on margins, distolateral surface, and mesial surface. Endite tapered distally and subequal to first segment of exopod. Exopod with two segments: proximal segment narrow, margins parallel, with plumose setae; distal segment spatulate, longer than wide, broadest medially, margins and mesioventral surface with long plumose setae. Endopod flattened and elongate, reaching twothirds to distal end of proximal exopodal segment; plumose setae on margins and median of lateral surface.
Maxilliped II (fig. 96I) dactylus evenly rounded, length subequal to width, with thick simple setae distally and on distolateral surface. Propodus two times wider than long, slightly produced at dorsodistal angle, with plumose setae on dorsal margin and patch of long simple setae on dorsodistal and ventrodistal angles. Carpus not produced dorsodistally, approximately two times longer than wide; long simple setae on dorsal margin. Merus approximately three times longer than wide, margins parallel; with simple and plumose setae on margins. Basisischium incompletely fused with plumose setae on margins. Exopod onehalf longer than merus, flagellum with one elongate article, approximately as long as carpus.
Maxilliped III (fig. 96J) dactylus oblong with rounded tip; long plumose setae on margins and lateral surface. Propodus dorsodistally inflated, with longitudinal median row of plumose setae on lateral surface; margins with plumose setae. Carpus produced onto propodus approximately onethird length of propodus; lateral surface with two rows of plumose setae; plumose setae on margins. Merus inflated, unarmed, with plumose setae on margins and few scattered small areas on lateral surface. Basisischium incompletely fused, with weak crista dentata of one or two teeth. Exopod twosegmented: proximal segment small; distal segment styliform, taper ing, approximately onethird length of merus; with plumose setae on margins; without flagellum.
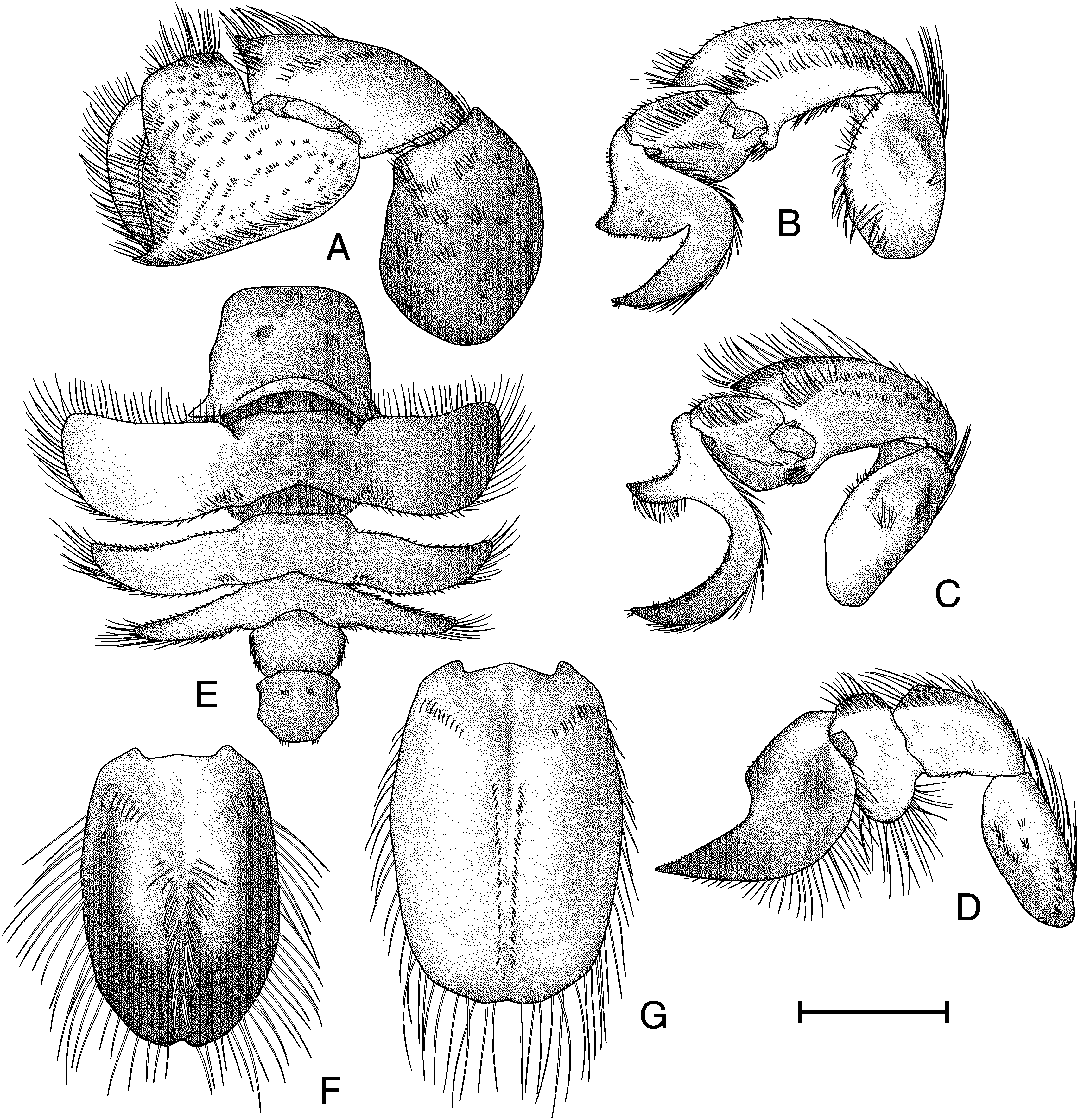
Pereopod I (fig. 97A) dactylus curved and tapering; lateral and mesial surfaces smooth; dorsal margin with long plumose setae; ventral margin with short simple setae. Propodal lateral surface with numerous short, trans verse rows of setose rugae; dorsal margin unarmed; ventral margin distally produced into acute spine; cutting edge lacking teeth, lined with long plumose setae; dorsal margin with long plumose setae, ventral margin with short simple setae. Carpus with dorsodistal angle produced into strong corneoustipped spine; dorsal margin with short transverse grooves behind spine; dorsal and distal margins with long plumose setae; lateral surface with small distal rugose area, few transverse setose ridges on distal half of surface; mesial surface smooth, with medial transverse row of setae, margins with long plumose setae. Merus unarmed; lateral surface with scattered transverse rows of long plumose setae, distal margin with long plumose setae; mesial surface with few scattered setae; fully calcified. Basisischium incompletely fused, unarmed. Coxa unarmed.
Pereopod II (fig. 97B) dactylus smooth; base to heel strongly and smoothly concave, heel produced, broad and subacute, heel to tip with narrow, acute indent, tip acute, tip to base broadly convex; lateral surface smooth, with several small tufts of short setae in generally straight line across medioproximal surface, several widely spaced submarginal tufts of short setae dorsodistally; mesial surface smooth, ventral margin with long plumose setae, dorsal margin with short simple setae, with patch of long plumose setae at base. Propodal dorsal surface smooth, with ventral margin inflated and rounded; oblique row of long plumose setae on distal margin of lateral surface; distal and ventral margins with long plumose setae; dorsolateral surface as narrow, oblique, flattened shelf, with short setae on dorsal margin and long plumose setae on ventral margin; mesial surface with elevated, curved, setose ridge from ventral junction with dactylus almost to ventral proximal junction with carpus. Carpus strongly produced and rounded dorsodistally, dorsal margin smooth; lateral surface smooth, produced area smooth, irregular, interrupted row of rugae and submarginal elevated ridge ventrally, rugae and ridge with long plumose setae; dorsodistal margin with long plumose setae, proximodorsal and ventral margins with short plumose setae; mesial surface smooth, with row of long plumose setae distally and subdorsally. Merus with large median decalcified window covering nearly all of lateral surface, long plumose setae on dorsodistal and ventral margins, few scattered long plumose setae on surface; mesial surface nearly smooth, with two long rows of setae. Basisischium incompletely fused and unarmed. Coxa unarmed.
Pereopod III (fig. 97C) dactylus with base to heel deeply concave, heel narrow and acutely produced, heel to tip with broadly concave indent and small concave region at midpoint of proximal margin, tip acute, tip to base smoothly convex; lateral surface smooth, with several small tufts of short setae in generally straight line across medioproximal surface, dorsodistal margin with tufts of short setae; ventral margin with long plumose setae, dorsal margin with short simple and plumose setae; mesial surface smooth, with plumose setae proximally at junction with propodus. Propodus not inflat ed dorsoventrally; lateral surface smooth, with long plumose setae in oblique row, simple setae on dorsal margin, plumose setae on ventral margin; dorsolateral surface narrow, oblique, flattened, with sparse setose mat; mesial surface smooth. Carpus produced dorsodistally, exceeding proximal margin of propodus by onethird length of propodus; tip subacute, dorsolateral margin unarmed; lateral surface slightly rugose dorsodistally, with mat of short setae and two interrupted rows of setae ventrally; mesial surface smooth, with long plumose setae on distal margin and in oblique row on surface. Merus smooth, with large decalcified window covering nearly half of lateral surface medially; dorsal and ventral margins unarmed, dorsodistal and ventrodistal margins with long plumose setae; mesial surface smooth. Basisischium incompletely fused and unarmed. Coxa unarmed. Female with large gonopore on anterior mesial margin of coxa, surround ed with short plumose setae; male without pore.
Pereopod IV (fig. 97D) dactylus with base to tip straight proximally, with subquadrate heel and broadly rounded, strongly concave indent and almost straight from indent to tip, tip acute, tip to base concave distally to convex proximally; lateral surface smooth, ventral margin with long plumose setae, dorsal margin with short simple setae; mesial sur face with dorsal decalcified region, demarcated ventrally by longitudinal elevated ridge with row of short setae; with setose punctations ventral to decalcified window. Propodus expanded dorsally and ventrally; ventral expansion reaching ventral margin of dactylus, ventral margin with long plumose setae; dorsal expansion with row of long plumose setae dorsally, oblique area with mat of short simple setae; lateral surface smooth, mesial surface smooth, with distoventral area of few patches of long plumose setae. Carpus slightly produced dorsodistally; ventral fourfifths of lateral surface and mesial surface smooth, dorsodistal onefifth of lateral surface with mat of short setae; dorsal margin with short simple and long plumose setae; ventral margin with short simple setae; mesial surface decalcified medially. Merus with scattered short transverse rows of setae on lateral surface, dorsal margin with long plumose setae; proximoventral half of mesial surface with large decalcified window. Basisischium incompletely fused and unarmed. Coxa unarmed.
Abdomen (fig. 97E) somite I wider than long, widest posteriorly; dorsal surface with anterior margin straight; posterior margin curved with elevated submarginal row of short setae; small transverse decalcified windows laterad of segment median. Somite II dorsal surface with irregular, submarginal, transverse ridge anteriorly; with small transverse, decalcified windows laterad of segment median just anterior to submarginal ridge; pleura expanded and directed anterolaterally; anterolateral margins angled, anterior and lateral margins with long plumose setae, posterolateral angle rounded, posterior margin with short setae; posteromesial angle with mat of short simple setae. Somite III similar to somite II, but narrower, shorter, anterior submarginal windows present; pleura thinner and shorter than on somite II, direct ed posterolaterally proximally and anterolaterally distally, with setae as in somite II; anterolateral angle subacute; dorsal surface obliquely flattened anterolaterally, with submarginal row of short setae. Somite IV similar to somite III, but thinner and shorter, anterior submarginal windows present; pleura thinner and shorter than on somite III, directed posterolaterally; dorsal surface obliquely flattened anterolaterally; lateral and posterior margins with long plumose setae, anterior margin with short simple setae. Somite V wider than somite IV, anterior submarginal windows present; lateral margins with plumose setae; pleura absent. Somite VI slightly broader than somite V, anterior submarginal windows present; dorsal surface with short transverse rows of setae laterad of midline and on posterior margin; pleura absent.
Females with uniramous, paired pleopods on somites II–V; males without pleopods.
Telson of male (fig. 97F) ovate and elongated, length greater than width, distal tip rounded, with median indentation; thickly calcified medially, inflated dorsally; distal twothirds with lateral decalcified region; median longitudinal groove extending onehalf length, row of long, thick, simple setae of either side of median groove beginning at median and continuing to distal margin of calcified area; proximolateral angles with long simple setae; margins with long simple setae. Telson of female (fig. 97G) similar to male, but more truncate distally and with smaller median indentation, dorsal surface evenly calcified; median groove similar to male, but with shorter setae; proximolateral angle with few short simple setae, margins with long simple setae.
DISTRIBUTION: From southern Japan southward in a narrow band to Western Australia and Victoria, Australia, in up to 45.7 m depth.
MAXIMUM SIZE: Males: 14.4 mm cl; females: 17.0 mm cl.
TYPE SPECIMENS: RMNH ex 32052 (holotype), RMNH 32052 (allotype); AM P5340 (2 paratypes), AM P7029 (paratype), BMNH,1932.11.30.65 (paratype), BMNH 1937.6.1.8 (paratype), MCZ 9625 (paratype), MOV J14552 View Materials (paratype), MOV J40194 View Materials (paratype), MOV J44728 View Materials (paratype), MOV J44732 View Materials (paratype), NMCR 1151b (paratype), NTOU (3 paratypes), NTOU (2 paratypes), QM W143 (paratype), QM W489 (paratype), QM W873 (paratype), QM W2221 (2 paratypes), USNM 59074 (paratype), WAM 20667 (3 paratypes), WAM 23386 (5 paratypes), WAM 23389 (6 paratypes), WAM 23391 (paratype), WAM 23395 (paratype), WAM 23396 (paratype), ZLKU 5125–5126 (2 paratypes), ZLKU 7438 (paratype), ZLKU 7511 (paratype), ZRC 1995.571 (paratype) .
TYPE LOCALITY: MikawaIsshiki, Aichi Prefecture, Honshu, Japan.
ETYMOLOGY: This species is named after Matt Groening, cartoonist and creator of the television program ‘‘The Simpsons,’’ to hon or his extensive promotion of crustacean issues in the popular media. ‘‘The Simpsons’’ has exposed people to a diversity of crustacean species, including Lisa’s hermit crab, Patty and Selma’s hermit crab cleaning techinques, and, of course, ‘‘Pinchy’’ the lobster. The specific name is pronounced ‘‘grayningi.’’
REMARKS: This species is more common in the Philippines than indicated by the above material examined. Serène and Umali (1965) cited 54 specimens from various Philippine locations (as A. steinitzi ). It is possible that some of Serène and Umali’s (1965) material is not this species, but the identification of their material is based on their descriptions and illustrations and a single specimen cited by them that was available for examination ( NMCR 1151b). Curiously, Serène and Umali (1965) stated that all of their specimens were males. However, their ‘‘female’’ of A. thurstoni (pl. 5, fig. 1a) is clearly a male, so they may have been in error regarding other specimens as well. The record of Kikuchi (1932) from Toyama Bay, Japan, is probably this species, the only Albunea known with certainty from that location.
One ♀ specimen from Kaoasiung, Taiwan ( NTOU) is host to an undescribed species of Albunione Markham and Boyko ( Isopoda : Bopyridae ).
This species belongs to the ‘‘ holthuisi group’’ of species and is the sistertaxon to A. holthuisi .
Albunea symmysta ( Linnaeus, 1758) Figures 98 View Fig , 99 View Fig
Cancer Symmysta Linnaeus, 1758: 630 View in CoL View Cited Treatment . – Linnaeus, 1764: 453.
Cancer symnista [sic]: Linnaeus, 1767: 1053.
Cancer Gymnista View in CoL [sic]: Houttuyn, 1769: 422.
Cancer gymnista [sic]: Statius Müller, 1775: 1128.
Hippa symnista [sic]: Fabricius, 1787: 329 (part). – Fabricius, 1793: 474.
Cancer Symnista View in CoL [sic]: de Villers, 1789: 157–158
(part). – Gmelin and Linne´, 1790: 2994. – Griffith and Pidgeon, 1833: 178.
Cancer dorsipes: Herbst, 1791: 5–8 , pl. 22, fig. 2. – Herbst, 1796: 197–198, pl. 45, figs. 1–7 (not Notopus dorsipes ( Fabricius, 1793)) View in CoL .
Albunea symnista [sic]: Weber, 1795: 94. – Fabricius, 1798: 397. – Herbst, 1804: 31 (list). – Latreille, 1806: 44. – Lamarck, 1818: 224. – Desmarest, 1823: 283, unnumbered pl., fig. 3. – Desmarest, 1825: 173, pl. 29, fig. 3. – Guérin Méneville, 1829: 12 (list), pl. 15, fig. 1. – Latreille, 1831: 56. – Brewster, 1832: 234. – H. Milne Edwards, 1837a: 111–112, explanation for pl. 42, pl. 42, fig. 3. – H. Milne Edwards, 1837b: 203–204, pl. 42, fig. 3. – H. Milne Edwards, 1840: 111–112. – White, 1847: 57. – de Haan, 1849: viii, pl. Q. – Lucas, 1853: 45–47, pl. 1, fig. 8. – Heller, 1863: 153 (part). – Heller, 1865: 72. – Chenu and Desmarest, 1877: 32. – Miers, 1878: 326–327. – Lucas, 1881: 54–55 *. – Albert, 1883: 523–524, pl. 31, figs. 1, 6, 17. – de Man, 1887: 425. – F. Müller, 1890: 472. – Ortmann, 1892: 536. – Henderson, 1893: 338, 409*. – Stebbing, 1893: 152 (list). – Southwell, 1910: 183. – Boschma, 1931: 351, 354*. – Boschma, 1937: 204 *. – Menon, 1937: 10–15, figs. 46–82. – Gordon, 1938: 187 (part), figs. 1e, 3f, 4c*. – Bouvier, 1940: 181, pl. 6, fig. 1. – Gravely, 1941: 75, 104, fig. 27–1. – Gurney, 1942: 263, fig. 110k. – Boschma, 1955: 14, 57, 65, 67*. – Sarojini, 1962: 191, fig. 1j. – Serène and Umali, 1965: 90–95 (part), pl. 1, fig. 1, pl. 2, fig. 1, pl. 3, fig. 1, 1a, pl. 4, fig. 1, textfigs. 1a, 2a, 3, 4a, 5a, b, 6a, b, b̍*. – Thomassin, 1969: 138–140, pl. 1, textfig. 3a. – Boonruang and Phasuk, 1975: 1–2, 8, 15, 17, fig. 4. – Anantaraman and Subramoniam, 1976: 192–199. – Naiyanetr, 1978: 333, fig. 1. – Naiyanetr, 1980: 22. – Panneerselvam and Subramoniam, 1983: 1–8, figs. 1–8. – Subramoniam, 1984: 78–94, figs. 1a, 2–14. – Coêlho and Calado, 1987: 43, table 1. – Seridji, 1988: 1298. – Subramoniam, 1993: 133, 152–155, 158, 197–198, figs. 14, 15. – Calado, 1997a: 17. – Naiyanetr, 1998: 51. – K. Sakai, 1999: 9, pl. 1, fig. d.
albunea symnista [sic]: Latreille, 1803: 172, pl. 51, fig. 4.
Albunea dorsipes: Herbst, 1804: 31 (list) (not Notopus dorsipes ( Fabricius, 1793)) View in CoL .
Cancer lymnista [sic]: Froriep, 1806: 183 (list).
Albunea Symniste View in CoL [sic]: Duméril, 1816: 431.
Albunaea [sic] symnista [sic]: Stimpson, 1858: 230 (list).
Albunea View in CoL ( symnista ): Claus, 1885: 69, 108, pl. 6, fig. 52. – Claus, 1886: 69, 108, pl. 6, fig. 52.
Albunea symmysta: Ortmann, 1896: 224 View in CoL . – Ortmann, 1901: 1276, pl. 72, fig. 3, pl. 93, fig. 4, pl. 104, fig. 2. – Holthuis, 1956: 238. – ICZN, 1958: 234. – Subramoniam and Panneerselvam, 1985: 226–227. – Boyko and Harvey, 1999: 391, 396, 400 (list), 401 (key)*. – Boyko, 1999: 145 (list).
Cancer symmysta: Holthuis, 1956: 237–238 . – ICZN, 1958: 213, 215, 233–234. – Melville and Smith, 1987: 298. Albunea Symnista View in CoL [sic]: Seridji, 1988: 1298.
Albunea symnysta [sic]: Chace and Kensley, 1992: 446, fig. 2n.
Cancer symmista [sic]: Calado, 1995: 71.
‘‘ Albunea View in CoL sp. n. ‘‘ Calado, 1995: 76–78, pl. 4, fig. l, pl. 5, fig. l, pl. 23, fig. a, pl. 24, figs. a–e*.
Albunea symnestra [sic]: Dexter, 1996: 12.
Albunea symmista [sic]: Fransen et al., 1997: 161 *. – Richmond, 1997: 214, unnumberef fig. on p. 215.
Albunea edsoni Calado, 1997a: 18–21 View in CoL , figs. 1, 2* (NEW SYNONYMY).
? Albunea oxyophthalma: Southwell, 1910: 184 View in CoL (not Albunea oxyophthalma Miers, 1878 View in CoL = A. paretii Guérin Méneville, 1853 View in CoL ).
? AIbunea [sic] symnista [sic]: Menon, 1937: 10.
? Albunea View in CoL [sp.] Menon, 1943: 331–332.
? Albunea symnista [sic]: Wang, 1989: 39. – Sun and Wang, 1996: 31 (list) (? = Albunea sp. indet.).
not Hippa symnista [sic]: Fabricius, 1787: 329 (part) (= Corystes cassivelaunus ( Pennant, 1777)) View in CoL .
not Cancer Symnista View in CoL [sic]: de Villers, 1789: 157– 158 (part) (= Corystes cassivelaunus ( Pennant, 1777)) View in CoL .
not Albunea symnista [sic]: RafinesqueSchmaltz, 1814: 20. – Lucas, 1849a: 27–28 *. – Lucas, 1849b: pl. 3, fig. 2*. – Heller, 1863: 153 (part). – Barrois, 1888: 18–19, 75, 82, 89, 93–94. – Bolivar, 1892: 128 (list). – Ferrer Aledo, 1914: 68. – Miranda y Rivera, 1933a: 22. – Miranda y Rivera, 1933b: 1 (list) (= Albunea carabus ( Linnaeus, 1758)) View in CoL .
not Albunea symnista [sic]: Brulle´, 1836–1844: 17. – Ozorio, 1888: 186 (= Albunea elegans A. Milne Edwards and Bouvier, 1898 View in CoL ).
not Albunea symnista [sic]: Gibbes, 1850a: 24 * (= Albunea paretii Guérin Méneville, 1853 View in CoL ).
not Albunea symnista [sic]: Gibbes, 1850b: 187 (= Albunea catherinae View in CoL , n. sp.).
not Albunea symnista [sic]: A. Milne Edwards, 1862: F–12* (= Albunea speciosa Dana, 1852 View in CoL ).
not Albanea [sic] symnista [sic]: Hoffman, 1874: 42 (list) (= Albunea speciosa Dana, 1852 View in CoL ).
not Albunea symnista [sic]: Cano, 1889a: 95, 104. – Cano, 1889b: 263. (= Albunea steinitzi Holthuis, 1958 View in CoL ).
not Albunea symmysta: Nobili, 1906: 142–143 View in CoL *. – Ramadan, 1936: 3 (list) (= Albunea steinitzi Holthuis, 1958 View in CoL ).
not Albunea symnista [sic]: Rathbun, 1924: 29 (= Albunea sp. indet.).
not Albunea symnista [sic]: Nakazawa, 1927: 1051, fig. 2025. – Nakazawa et al., 1949: 741, fig. 2144. – Nakazawa et al., 1951: 741, fig. 2144 (=? Paralbunea dayriti (Serène and Umali, 1965)) View in CoL .
not Albunea symnista [sic]: Kikuchi, 1932: 10 (= Albunea View in CoL ? groeningi View in CoL , n. sp.).
not Albunea symnista [sic]: T. Sakai, 1935: 61 (= Albunea sp. indet.).
not Albunea symnista [sic]: Gordon, 1938: 187 (part)*. – Serène and Umali, 1965: 90–95 (part)*. – Miyake, 1991: 157, pl. 53, fig. 2. – Asakura, 1995: 376, fig. 21–285 (= Albunea occultus , n. sp.).
not Albunea symnista [sic]: Gordon, 1938: 187 (part)*. – Miyake, 1978: 152–154, figs. 59, 60a* (= Albunea groeningi View in CoL , n. sp.).
not Albunea symnista [sic]: Gordon, 1938: 187 (part)* (= Albunea bulla View in CoL , n. sp.).
not A. symnista [sic]: Ward, 1942: 52 (list), 63 (=? Albunea holthuisi Boyko and Harvey, 1999 View in CoL ).
not Albunea symnista [sic]: Barnard, 1950: 405– 406. – Kensley, 1981: 35 (list) (= Albunea sp. indet.).
not Albunea symnista [sic]: Kikuchi, 1959: 49 (list). – Kikuchi, 1961: 5 (list) (= Albunea sp. indet.).
not Albunca [sic] symnista [sic]: Kikuchi, 1961: 5 (list) (= Albunea sp. indet.).
not Albunea symmysta: Miyake, 1961: 12 View in CoL .— Miyake et al., 1962: 125 (part) (=? Paralbunea dayriti (Serène and Umali, 1965)) View in CoL .
not Albunea symmysta: Miyake et al., 1962: 125 View in CoL (part, Toyama Bay material) (= Albunea groeningi View in CoL , n. sp.).
not Albunea symmista [sic]: Miyake, 1965: 651 * (= Albunea occultus , n. sp.).
not Albunea symmista [sic]: Miyake, 1965: fig. 1111 (=? Paralbunea dayriti (Serène and Umali, 1965)) View in CoL .
not Albunea symnista [sic]: Serène, 1977: 47. – Calado, 1995: 71–73, pl. 4, fig. i, pl. 5, fig. h, pl. 21, figs. a, b, pl. 22, figs. a–f* (= Albunea microps Miers, 1878 View in CoL ).
not Albunea symmysta: Kikuchi and Miyake, 1978: 31 View in CoL (list) (= Albunea sp. indet.)
not Albunea symmysta: Nurul Huda et al., 1989: 88–89 View in CoL (= Albunea View in CoL ? thurstoni Henderson, 1893 View in CoL ).
MATERIAL EXAMINED: India: ‘‘ Indian Seas,’’ coll. unknown: 1 Ƌ, 13.6 mm cl, 2 ♀, 19.0– 19.8 mm cl ( BMNH 1883.25 ) ; ‘‘ Ganjam ,’’ coll. unknown: 1 ♀, 15.0 mm cl ( BMNH 1889.6.17.151) ; Colachel , coll. un known: 1 Ƌ, 14.1 mm cl ( BMNH 1903.4.6.210) ; Madras , coll. J. R. Henderson: 1 Ƌ, 16.2 mm cl, 7 ♀, 14.2–21.9 mm cl ( BMNH 1892.7.15.125–132) ; Madras , coll. unknown: 3 ♀, 17.2–19.6 mm cl ( BMNH 1903.4.6.211–212) ; [ Madras ], coll. unknown, 1 larva, 2.8 mm cl ( BMNH 1949.12 .2.544 ex Menon via Gurney) ; Madras , coll. Museum Wien: 1 Ƌ, 14.8 mm cl, neotype ( ZMO F17538 View Materials ) ; Pondicherry, 1881, coll. M. Maindron: 1 ♀, 18.0 mm cl (MNHNHi 115) ; Pondicherry, 1881, coll. M. Maindron: 3 Ƌ, 12.5–13.4 mm cl (MNHNHi 116); Pondicherry, Jan. 1881, coll. M. Maindron: 2 ♀, 15.3–18.0 mm cl (MNHNHi 16) ; Pondicherry, Nov. 28, 1942, coll. unknown: 1 ♀, 19.5 mm cl (MNHNHi 15) ; Pondicherry, coll. M. Leschenault: 1 Ƌ, 17.4 mm cl, 1 ♀, 17.8 mm cl (MNHNHi 182) ; Pondicherry, coll. M. Leschenault: 1 ♀, 14.7 mm cl (MNHNHi 183) ; Pondicherry, coll. M. Leschenault: 1 ♀, 16.4 mm cl (MNHNHi 184) ; Pondicherry, coll. M. Leschenault: 1 Ƌ, 13.0 mm cl (MNHNHi 185); [Pondicherry], coll. M. Leschenault: 1 Ƌ, 14.1 mm cl (MNHNHi 186); Pondicherry, Aug. 1901, coll. M. Maindron: 25 Ƌ, 10.6– 15.5 mm cl, 16 ♀, 11.0 – 20.8 mm cl (MNHNHi 196) ; Pondicherry, Bay of Bengal , 11°59̍N, 79°50̍E, 1901, coll. [M. Maidron]: 1 Ƌ, 14.6 mm cl (AM P5404) ; Pondicherry, 1901, coll. [M. Maidron]: 1 Ƌ, 15.5 mm cl (AM P5405 ex MNHN) ; Pondicherry, 1901, coll. [M. Maidron]: 1 Ƌ, 13.0 mm cl (AM P5406 ex MNHN) .
Sri Lanka (Ceylon): ‘‘ Ceylon,’’ coll. E. E. Green: 2 Ƌ, 15.3–15.9 mm cl, 1 ♀, 16.2 mm cl ( BMNH 1904.11.28.4–6) ; ‘‘ Beutkofe, SW Ceylon,’’ Aug. 2, 1909, coll. G. Dünker: 1 ♀, 17.7 mm cl ( ZMH K–32584) .
Indonesia: Padang , Sumatra, Oct. 14, 1828 – Nov. 10, 1829, coll. S. Müller: 2 ♀, 15.0–16.0 mm cl ( RMNH 23029 About RMNH ) ; Benkoelen , Sumatra, Nov. 1925, coll. H. C. Kellers: 1 ♀, 19.1 mm cl ( USNM 68615 About USNM ) ; ‘‘ Buitenzorg ,’’ Java, June 1906, coll. H. Jensen: 16 Ƌ, 10.2–14.1 mm cl, 24 ♀, 11.2–19.2 mm cl ( ZMUC 2713 View Materials ) ; ‘‘ Moluccas ,’’ 1841–1843, coll. E. A. Forsten: 1 Ƌ, 15.5 mm cl, 1 ♀, 13.5 mm cl ( RMNH 23030 About RMNH ) ; ‘‘ Moluccas ,’’ coll. unknown: 8+ unsexable specimens, 12.5–15.3 mm cl ( RMNH 853 About RMNH ) .
Thailand: Phuket Island, July 1970, coll. unknown: 1 ♀, 19.4 mm cl ( CASIZ 109242).
Philippines: Padada Beach, Gulf of Davao, Sept. 1939, coll. G. R. Oesch: 1 oviger, 12.9 mm cl ( AMNH 10393 About AMNH ) ; Morong , Bataan, March 1, 1999, coll. J. D. Williams: 2 Ƌ, 8.9–12.6 mm cl, 1 juvenile, 5.2 mm cl ( AMNH 18092 About AMNH ) ; Morong , Bataan, Feb. 20, 1999, coll. J. D. Williams: 1 molt, 7.1 mm cl ( AMNH 18093 About AMNH ) ; Levgayen Gulf , 500 ft (= 151.5 m), May 1909, coll. ‘‘ Albatross’ ’: 1 ♀, 14.8 mm cl ( USNM 68613 About USNM ) ; Calapan, Mindoro , Jan. 6, 1933, coll. P. de Mesa: 1 Ƌ, 12.1 mm cl ( MCZ 19654) ; Calapan, Mindoro , coll. P. de Mesa: 1 Ƌ, 8.2 mm cl ( MCZ 9615 About MCZ ) .
Australia: Queensland: 15°28̍S, 145°15̍E, Cooktown , June 1, 1985, coll. S. W. Gunn: 1 Ƌ, 9.7 mm cl ( MOV J44730 View Materials ) ; Cardwell, Port Douglas , Jan. 20, 1972, coll. E. M. Grant: 1 ♀, 15.1 mm cl ( QM W3848 ) ; New South Wales: Lord Howe Island , coll. unknown: 1 ♀, 17.8 mm cl, holotype of A. edsoni ( BMNH 1912.11.22.106) .
Limited Data: Sta. 1679, Mission de l’Indochine, 1912, coll. A. Kreuff: 4 Ƌ, 12.2–13.2 mm cl, 2 ♀, 13.6–15.5 mm cl (MNHNHi 86) ; ‘‘ Asia ,’’ pre–1866, coll. L. de Jeude: 1 ♀, 18.7 mm cl ( MOV J14551 View Materials ) ; ‘‘ Asia ,’’ coll. L. de Jeude: 1 Ƌ, 11.9 mm cl ( MOV J44726 View Materials ) ; ‘‘ Yanaon, Indes Orientales ,’’ coll. unknown: 1 ♀, 18.6 mm cl ( MNHN Hi 187) .
No Data: 1 Ƌ, 12.4 mm cl (MNHNHi 17); 1 ♀, 17.0 mm cl ( MOV J14554 View Materials ) .
DIAGNOSIS: Carapace wider than long, covered with lightly setose grooves. Anterior margin with 11–17 spines on either side of ocular sinus. Setal field with narrow lateral elements and concave anterior margin. CG1 with separate posterior lateral elements; CG4 with two short, anteriorly displaced medial elements between longer supralateral elements of CG4; CG5 of two triangular elements; CG6 and CG7 separate; CG8 broken; CG11 absent. Rostrum present, not reaching posterior margin of ocular plate. Ocular plate triangular. Distal peduncular segments dorsoventrally flattened and triangular, tapering at tip, approximated along mesial margins at base, lateral margins broadly convex, mesial margins straight. Cornea at tip. Dactylus of pereopod II with heel produced, tapered, and subacute. Dactylus of pereopod III with heel tapering, projecting, acute. Dactylus of pereopod IV sinuous from base to tip, with produced subacute heel and deep indent. Telson of male broadly triangular, laterally produced, length subequal to width, with broadly rounded tip; thickly calcified medially, inflated dorsally; distal twothirds with lateral decalcified region, median row of thin setae. Telson of female flattened, ovate, and evenly calcified with slightly produced tip.
DESCRIPTION: Carapace (fig. 98A) slightly wider than long. Anterior margin slightly concave on either side of ocular sinus, becoming convex laterally, with 11–17 large spines (n = 9) along length. Rostrum as small acute tooth, not reaching proximal margin of ocular plate. Ocular sinus smoothly concave, with three or four small spinules. Frontal region smooth; setal field narrow anteriorly and posteriorly; posterior lateral elements reduced to narrow bands of setae. CG1 parallel to anterior margin of carapace, sinuous, strongly crenulate, divided into medial fragment and curved, posteriorly displaced lateral elements. Mesogastric region smooth; CG2 absent; CG3 broken into five or six short elements between posterior lateral elements of CG1, medial elements anteriorly displaced; CG4 with two short, anteriorly displaced medial elements between longer supralateral elements of CG4. Hepatic region smooth, with oblique setose groove at median of lateral margin. Epibranchial region generally triangular, smooth; posterolateral margin without rows of setae. Metagastric region smooth; CG5 present as two triangular elements. CG6 strongly crenulate, strongly anteriorly concave medially and sloping out to anteriorly convex lateral thirds. CG7 oblique, not reaching lateral margins of median segment of CG6. Cardiac region smooth; CG8 present as two to four short medial elements. CG9 present as two short lateral grooves with gap at midline. CG10 absent. CG11 absent. PostCG11 element absent. Branchial region with numerous short, transverse rows of setae in anterior half. Posterior margin deeply and evenly convex, with submarginal groove reaching twothirds up margin of posterior concavity. Branchiostegite with short anterior submarginal spine; anterior region with scattered, short, transverse lines ventral to linea anomurica; with many short rows of setae and sparsely covered with long plumose setae ventrally; posterior region membranous, with numerous irregular fragments and sparsely covered with long plumose setae.
Ocular plate (fig. 98B) triangular, with deep median indentation; median peduncular segments present as small ovate calcified areas lateral to ocular plate. Distal peduncular segments elongate, subtriangular, with strongly convex lateral and straight mesial margins, cornea covering lateral portion of distal tip; lateral margins with notch onefourth distal from base; mesial margins approximated at base, with long plumose setae; tuft of plumose setae at proximolateral ventral angles and ventromedial oblique row of plumose setae extending from tuft to threefourths length of segment.
Antennule (fig. 98C) with segment III narrow proximally, expanding distally to two times proximal width; with plumose setae on dorsal and ventral margins and sparsely scattered on lateral surface; dorsal exopodal flagellum with 121–131 articles (n = 6), long plumose setae on dorsal and ventral margins; ventral endopodal flagellum with three to five articles (n = 6), plumose setae on dorsal and ventral margins. Segment II medially inflated in dorsal view, with plumose setae on dorsal and ventral margins and scattered on ventrolateral third of surface. Segment I wid er than long, unarmed; dorsal third of lateral surface rugose with long plumose setae; long plumose setae on dorsal and ventral margins.
Antenna (fig. 98D) with segment V approximately two times longer than wide, with long plumose setae on dorsal and ventral margins and scattered on lateral surface; flagellum with seven articles (n = 6), long plumose setae on dorsal, ventral, and distal margins. Segment IV expanded distally, with long plumose setae on dorsal, ventral, and distal margins and row of setae on dorsolateral surface. Segment III with long plumose setae on dorsal and ventral margin and in short row on surface. Segment II short, widening distally, rugose, with plumose setae on margins and scattered on lateral surface; antennal acicle long, thin, and exceeding distal margin of segment IV by onesixth length of segment IV, with long plumose setae on dor sal margin. Segment I rounded proximally, flattened ventrolaterally, with long plumose setae on margins and in short row on surface rugae behind spine; lateral surface with acute spine dorsodistally, with low semicircular dorsolateral lobe ventrodistal to spine; segment with ventromesial antennal gland pore.
Mandible (fig. 98E) incisor process with three teeth; cutting edge with one tooth. Palp threesegmented, with plumose setae on margins and long, thick, simple setae arising from bend in second segment and on distal margin of terminal segment.
Maxillule (fig. 98F) distal endite proximally narrow, widening to inflated distal end, with thick simple setae on distal margin and thin simple setae on dorsal margin. Proximal endite with thick simple setae on distal margin. Endopodal external lobe truncate distally and curled under; internal lobe reduced with four thick setae at distolateral margin.
Maxilla (fig. 98G) exopod evenly rounded, with plumose setae along distal margin. Scaphognathite bluntly angled on posterior lobe, with plumose setae.
Maxilliped I (fig. 98H) epipod with plumose setae on margins, distolateral surface, and mesial surface. Endite tapered distally and subequal to first segment of exopod. Exopod with two segments; proximal segment narrow, margins parallel with plumose setae; distal segment spatulate, longer than wide, broadest medially, margins and mesioventral surface with long plumose setae. Endopod flattened and elongate, reaching twothirds to distal end of proximal exopodal segment; plumose setae on margins and median of lateral surface.
Maxilliped II (fig. 98I) dactylus evenly rounded, length slightly greater than width, with thick simple setae distally and on distolateral surface. Propodus 1.5 times wider than long, slightly produced at dorsodistal angle, with plumose setae on dorsal margin and patch of long simple setae on dorsodistal and ventrodistal angles. Carpus not produced dorsodistally, approximately two times longer than wide; long simple setae on dorsal margin. Merus approximately three times longer than wide, margins parallel; with simple and plumose setae on margins and scattered on surface. Basisischium incompletely fused with plumose setae on margins. Exo pod onethird longer than merus, flagellum with one elongate article, approximately as long as carpus.
Maxilliped III (fig. 98J) dactylus oblong with rounded tip; long plumose setae on margins and lateral surface. Propodus dorsodistally inflated, with longitudinal median row of plumose setae on lateral surface; margins with plumose setae. Carpus produced onto propodus almost onethird length of propodus; lateral surface with two rows of plumose setae on surface; plumose setae on margins. Merus inflated, unarmed, with plumose setae on margins and scattered on lateral surface. Basisischium incompletely fused, with weak crista dentata of three or four teeth. Exopod twosegmented: proximal segment small; distal segment styliform, tapering, approximately twofifths length of merus; with plumose setae on margins; without flagellum.
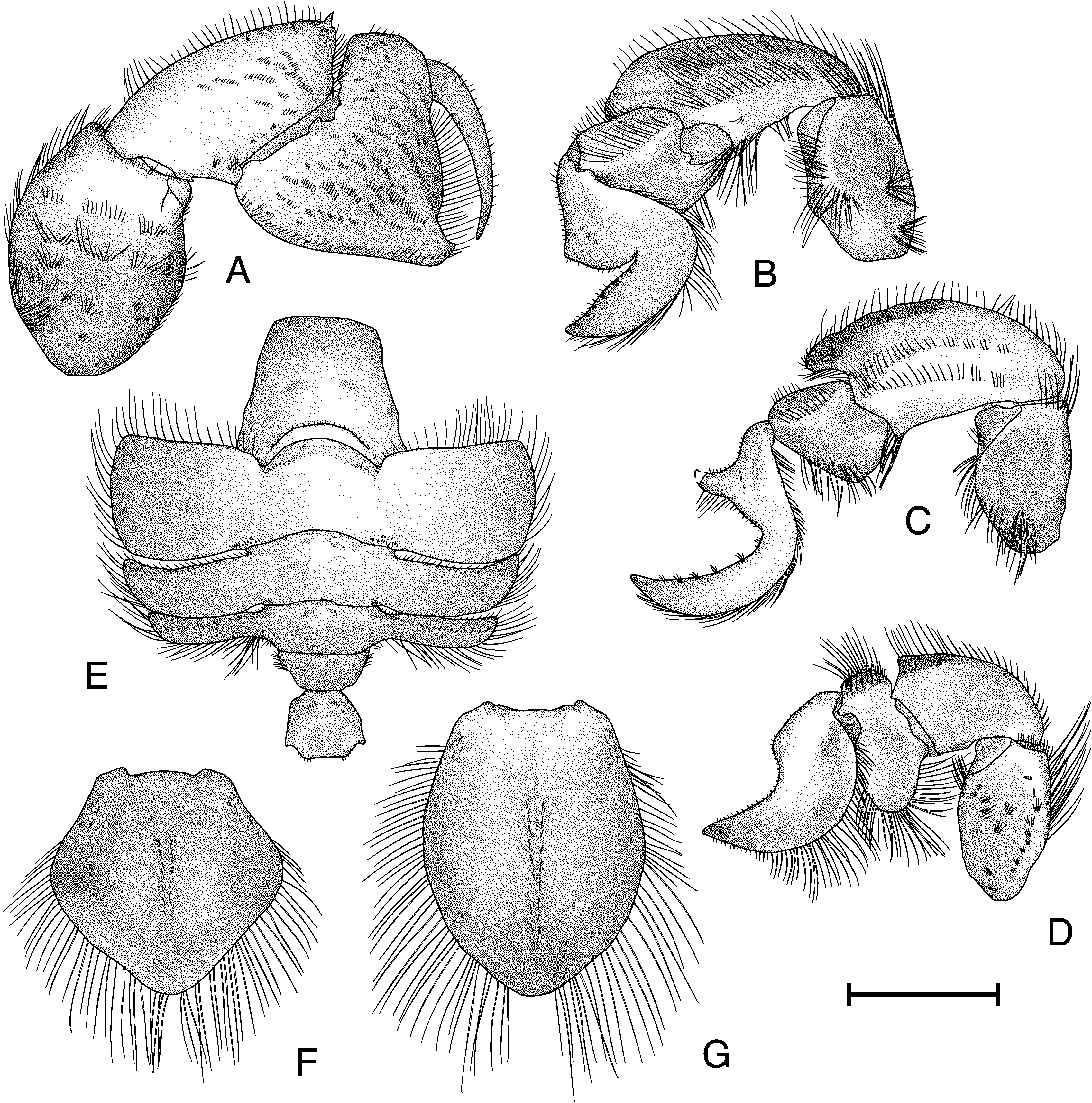
Pereopod I (fig. 99A) dactylus curved and tapering; lateral and mesial surfaces smooth; dorsal margin with long plumose setae; ventral margin with short simple setae. Propodal lateral surface with numerous short, transverse rows of setose rugae; dorsal margin unarmed; ventral margin distally produced into acute spine; cutting edge lacking teeth, lined with long plumose setae; dorsal margin with long plumose setae, ventral margin with short simple setae. Carpus with dorsodistal angle produced into strong corneoustipped spine; dorsal margin with short transverse grooves behind spine; dorsal and distal margins with long plumose setae; lateral surface with small distal rugose area, few transverse setose ridges on distal half of surface; mesial surface smooth, with medial transverse row of setae, margins with long plumose setae. Merus unarmed; lateral surface with scattered transverse rows of long plumose setae, margins with long plumose setae; mesial side with few scattered setae; fully calcified. Basisischium incompletely fused, unarmed. Coxa unarmed.
Pereopod II (fig. 99B) dactylus smooth; base to heel slightly concave, heel produced, broad and subacute, heel to tip with narrow, acute indent, tip acute, tip to base broadly convex; lateral surface smooth, with several small tufts of short setae in generally straight line across medioproximal surface, several widely spaced submarginal tufts of short se tae dorsodistally; mesial surface smooth, ventral margin with long plumose setae, dorsal margin with short simple setae, patch of long plumose setae at base. Propodal dorsal surface smooth, with ventral margin inflated and rounded; oblique row of long plumose setae on distal margin of lateral surface; distal and ventral margins with long plumose setae; dorsolateral surface as narrow, oblique, flattened shelf, with short setae on dorsal margin and long plumose setae on ventral margin; mesial surface with elevated, curved setose ridge from ventral junction with dactylus almost to ventral proximal junction with carpus. Carpus strongly produced and rounded dorsodistally, dorsal margin smooth; lateral surface smooth, produced area smooth, irregular, interrupted row of rugae and submarginal elevated ridge ventrally, rugae and ridge with long plumose setae; margins with short plumose setae; mesial surface smooth, with row of long plumose setae distally and subdorsally. Merus with large median decalcified window covering nearly all of lateral surface, with few scattered long plumose setae on surface and margins; mesial surface nearly smooth, with two long rows of setae. Basisischium incompletely fused and unarmed. Coxa unarmed.
Pereopod III (fig. 99C) dactylus with base to heel concave, heel narrow and acutely produced, heel to tip with broadly concave indent and small concave region at midpoint of proximal margin, tip acute, tip to base smoothly convex; lateral surface smooth, with several small tufts of short setae in roughly straight line across medioproximal surface, dorsodistal margin with tufts of short setae; ventral margin with long plumose setae, dorsal margin with short simple and plumose setae; mesial surface smooth, with plumose setae proximally at junction with propodus. Propodus not inflated dorsoventrally; lateral surface smooth, with long plumose setae in oblique row, simple setae on dorsal margin, plumose setae on ventral margin; dorsolateral surface narrow, oblique, flattened, with setose mat; mesial surface smooth. Carpus produced dorsodistally, exceeding proximal margin of propodus by onethird length of propodus; tip rounded, dorsolateral margin unarmed; lateral surface slightly rugose dorsodistally, with mat of short setae and two interrupted rows of setae ventrally; mesial surface smooth, with long plumose setae on distal margin and in oblique row on surface. Merus smooth, with large decalcified window covering nearly half of lateral surface medially; dorsal and ventral margins unarmed, with long plumose setae; distolateral margin with long plumose setae; mesial surface smooth. Basisischium incompletely fused and unarmed. Coxa unarmed. Female with large gonopore on anterior mesial margin of coxa, surrounded with short plumose setae; male with tiny pore located more mesially.
Pereopod IV (fig. 99D) dactylus with base to tip convex proximally, with broadly rounded, strongly concave indent and almost straight from indent to tip, tip acute, tip to base concave distally to convex proximally; lateral surface smooth, ventral margin with long plumose setae, dorsal margin with short simple setae; mesial surface with dorsal decalcified region, demarcated ventrally by longitudinal elevated ridge with row of short setae; with setose punctations ventral to decalcified window. Propodus expanded dorsally and ventrally; ventral expansion reaching ventral margin of dactylus, ventral margin with long plumose setae; dorsal expansion with row of long plumose setae dorsally, oblique area with mat of short simple setae; lateral surface smooth, mesial surface smooth, with distoventral area of few patches of long plumose setae. Carpus slightly produced dorsodistally; ventral fourfifths of lateral surface and mesial surface smooth, dorsodistal fifth of lateral surface with mat of short setae; dorsal margin with short simple and long plumose setae; ventral margin with short simple setae; mesial surface decalcified medially. Merus with scattered, short, transverse rows of setae on lateral surface, dorsal and ventrodistal margins with long plumose setae; proximoventral half of mesial surface with large decalcified window. Basisischium incompletely fused and unarmed. Coxa unarmed.
Abdomen (fig. 99E) somite I wider than long, widest posteriorly; dorsal surface with anterior margin straight; posterior margin curved, with elevated submarginal row of short setae; small transverse decalcified windows laterad of segment median. Somite II dorsal surface with irregular submarginal transverse ridge anteriorly; with small transverse decalcified windows laterad of segment median just anterior to submarginal ridge; pleura expanded and directed anterolaterally; anterolateral margins angled, anterior and lateral margins with long plumose setae, posterolateral angle rounded, posterior margin with short setae; posteromesial angle with mat of short simple setae. Somite III similar to somite II, but narrower, shorter, anterior submarginal windows present; pleura thinner and shorter than on somite II, directed anterolaterally, with setae as in somite II; anterolateral angle subacute; dorsal surface obliquely flattened anterolaterally, with submarginal row of short setae. Somite IV similar to somite III, but thinner and shorter, anterior submarginal windows present; dorsal surface with few short setae anterolaterally; pleura thinner and shorter than on somite III, directed anterolaterally; dorsal surface obliquely flattened anterolaterally; margins with long plumose setae. Somite V wider than somite IV, anterior submarginal windows present; lateral margins with plumose setae; pleura absent. Somite VI slightly broader than somite V, anterior submarginal windows present; dorsal surface with short transverse rows of setae laterad of midline; pleura absent.
Females with uniramous, paired pleopods on somites II–V; males without pleopods.
Telson of male (fig. 99F) broadly triangular, laterally produced, length subequal to width, with broadly rounded tip; thickly calcified medially, inflated dorsally; distal twothirds with lateral decalcified region; median longitudinal groove extending onehalf length, row of long simple setae of either side of median groove beginning at median and continuing almost to distal margin of calcified area; proximolateral angles with few long simple setae; margins with long simple setae. Telson of female (fig. 99G) flattened, ovate, and evenly calcified with slightly produced tip; median groove similar to male, setal row from midpoint of median groove to near distal margin of telson with simple setae subequal to those of male; proximolateral angle with few short setae, margins with long simple setae.
DISTRIBUTION: From the east coast of India throughout southeast Asia to the Philippines and Indonesia as far east as Java, in up to 151.5 m depth. Also Queensland and Lord Howe Island, Australia.
MAXIMUM SIZE: Males: 17.4 mm cl; females: 21.9 mm cl.
TYPE SPECIMENS: The type or types of Cancer symmysta are not extant and a neotype (Ƌ, 14.8 mm cl) is herein designated as ZMO F17538 View Materials . BMNH 1912.11.22.106 (holotype of A. edsoni ).
TYPE LOCALITIES: ‘‘Asia’’ was the only locality information given by Linnaeus (1758) for A. symmysta ; the neotype deisgnated herein ( ZMO F17538 View Materials ) was collected from Madras , India, which becomes the type locality of A. symmysta ; Lord Howe Island , New South Wales, Australia ( A. edsoni ) .
REMARKS: Albunea symmysta is no. 1565 on the ‘‘Official list of specific names in zo ology’’ ( ICZN, 1958) and is the type of the genus Albunea by subsequent designation of Holthuis (1956). This species is called ‘‘Jakalan quai’’ or ‘‘buffalo cicada’’ in Thailand, terms also used for the genus Hippa (Boonruang and Phasuk, 1975) .
Although Linnaeus (1758, 1767) gave the type locality as ‘‘Asia,’’ it should be noted that he also (1764) cited India as the ‘‘habitat’’ for this species, which is most likely where his specimens came from. Consequently, the neotype specimen ( ZMO F17538 View Materials ) is selected from an Indian locality. A neotype is needed to stabilize the identity of this species, as there have been numerous instances of confusion between other albuneid taxa and A. symmysta (see synonymy list above). The socalled holotype of Cancer dorsipes ‘‘Herbst’’ ( ZMB Herbst 2231), as cited by K. Sakai (1999), is merely Herbst’s (1804) specimen of A. symmysta . The species dorsipes was established in the genus Hippa by Fabricius in 1793.
The misspelling ‘‘ symnista ’’ was introduced by Linnaeus (1767) and unfortunately became the spelling used by most authors up to the present. It is hoped that the correct original spelling of this species will now be universally adopted.
Albunea symmysta is a model animal that has been used to study endocrinology, spermatophore formation, and breeding patterns (Panneerselvam and Subramoniam, 1983; Subramoniam, 1984; Subramoniam and Panneerselvam, 1985). Because of this, more is known about the biology of this species than any other albuneid, but not all published statements are accurate. As discussed for the family and genus, this species is neither blind nor a filterfeeder, as suggested by Subramoniam and Panneerselvam (1985).
This species breeds continuously along the Madras coast, with peaks in January and June, but the maximum percentage of ovigers is only 25% (Subramoniam and Panneerselvam, 1985). Sexual maturity of females was reported as 17 mm cl, and maximum size as 22 m cl (Subramoniam and Panneerselvam, 1985). However, an ovigerous ♀ of 12.9 mm cl is known from the Philippines ( AMNH 10393). The larval stages were described by Menon (1937) from Indian plankton samples, and there appear to be five zoeal stages. There are 12 gills along each side in the stage IV zoea and 13 in stage V zoea vs. 10 in an adult. Menon (1943) incorrectly suggested that this showed evidence of recapitulation of ancestral characters in the larval stages.
The androgenic gland of A. symmysta , as described by Sarojini (1962), is distinctive and unlike that of the pagurid hermit crab he studied, but the apparent overall variability in this gland within a single species makes its utility in phylogenetic studies questionable.
Albunea symmysta has been well studied with regards to its parasites. One 12.3 mm cl ♀ from Java ( ZMUC 2713) has an attached rhizocephalan parasite, Sacculina anceps Boschma, 1931 . This parasite is one of only a few species of ‘‘ Sacculina ’’ not found on brachyurans and may not properly belong to this genus (but see Boschma, 1955). Sacculina anceps was described from six specimens found on specimens of A. symmysta (ZMUC 2713) . A lectotype for S. anceps was selected by Boschma (1937) as ‘‘the best preserved of the two specimens of which longitudinal sections have been made’’. The specimen cited above is therefore a paralectotype of S. anceps . High levels of infection in Indian populations of A. symmysta by microphallid trematode metacercariae (Platyhelminthes) were reported by Anantaraman and Subramoniam (1976). The parasites were tentatively identified as Microphallus sp. and were found in the ovaries of female crabs, while male crabs were uninfected.
As A. symmysta is the only species known at present from either Sri Lanka or India, it seems likely that this is the taxon Southwell (1910: 184) collected from near Ceylon, in spite of his having also listed A. symmysta as a separate taxon ( Southwell, 1910: 183).
It is difficult to know what species Serène and Umali (1965) were dealing with from the Philippines, but at least some of their material was true A. symmysta . This can be seen in their illustrations of the inflated distal peduncular segments (textfig. 5a) and dactyli (textfig. 3), which match well with A. symmysta . Some of their material is clearly not A. symmysta , however, as can be seen in their discussion of the variable shape of the dactylus of pereopod IV. These other specimens are probably A. occultus , but need to be examined for confirmation.
Thomassin (1969) reported this species from Madagascar, but he had no specimens and relied on the personal communication of two individuals for his information. Howev er, the species is otherwise not known to occur in Madagascar and probably does not occur west of India. Because of this and numerous other misidentifications in the literature, Thomassin’s (1969: textfig. 11) distribution map for this species is not accurate. The citation of Richmond (1997) appears to be based on the same incorrect data from Thomassin (1969) and not on actual specimens from east Africa.
The identities of the dry specimens cited by Fransen et al. (1997) were confirmed by examination of sketches kindly supplied by Dr. Fransen.
Calado (1997a) described the species Albunea edsoni , based on a single ♀ specimen from Australia, but this specimen is actually a large example of A. symmysta . The main diagnostic feature given by Calado (1997a) was ‘‘17 teeth on the right frontal edge and 16 on the left.’’ As shown herein, the number of teeth on the frontal margin of Albunea species is only useful for separating species groups such as the ‘‘ microps group’’ from the ‘‘ symmysta group.’’ It is not a valid means of identifying species. Although not explicitly stated by Calado (1997a), the probable reason she described this new species was because the specimen was not conspecific with the single specimen of ‘‘ A. symmysta ’’ she examined ( Calado, 1995). I have examined that specimen ( BMNH 1956.1.14.20) and, as can be seen from Calado’s (1995: pl. 4, fig. i, pl. 5, fig. h, pl. 21, figs. a, b, pl. 22, figs. a–f) illustrations, it is actually a misidentified A. microps . Calado (1995, 1997a) apparently uncritically accepted the identification of this specimen on the label as written, and she incorrectly concluded that the Lord Howe Island specimen represented an undescribed species. This demonstrates the importance of studying large series of specimens in order to correctly understand intraspecific variation.
This species is closely related to A. occultus and these two species appear to be inter mediate in form between the ‘‘ holthuisi group’’ and the ‘‘ paretii group.’’

| RMNH |
National Museum of Natural History, Naturalis |
| NTOU |
Institute of Marine Biology, National Taiwan Ocean University |
| ZRC |
Zoological Reference Collection, National University of Singapore |
| WAM |
Western Australian Museum |
| QM |
Queensland Museum |
| ZMO |
Zoology Museum, Oxford University |
| MNHN |
Museum National d'Histoire Naturelle |
| ZMH |
Zoologisches Museum Hamburg |
| MCZ |
Museum of Comparative Zoology |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
| AMNH |
American Museum of Natural History |
| ZMUC |
Zoological Museum, University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Albunea groeningi
| BOYKO, CHRISTOPHER B. 2002 |
Albunea symmista
| Fransen, C. H. J. M. & L. B. Holthuis & J. P. H. M. Adema 1997: 161 |
| Richmond, M. D. 1997: 214 |
Albunea edsoni
| Calado, T. C. dos 1997: 21 |
Albunea symnestra
| Dexter, D. M. 1996: 12 |
Cancer symmista
| Calado, T. C. dos 1995: 71 |
Albunea
| Calado, T. C. dos 1995: 76 |
Albunea symnista
| Wang, F. 1989: 39 |
Albunea symmysta:
| Nurul Huda, K. M. & Q. Banu & B. Ahmed 1989: 89 |
Albunea symmysta: Kikuchi and Miyake, 1978: 31
| Miyake, S. 1978: 31 |
Albunea steinitzi: Serène and Umali, 1965: 97– 102
| Haig, J. 1974: 447 |
Albunea symmista
| Miyake, S. 1965: 651 |
Albunea symmysta:
| Miyake, S. & K. Sakai & S. Nishikawa 1962: 125 |
Albunea symmysta:
| Miyake, S. & K. Sakai & S. Nishikawa 1962: 125 |
Albunea symmysta: Miyake, 1961: 12
| Miyake, S. & K. Sakai & S. Nishikawa 1962: 125 |
| Miyake, S. 1961: 12 |
Albunea symnista
| Kikuchi, K. 1961: 5 |
| Kikuchi, K. 1959: 49 |
Cancer symmysta: Holthuis, 1956: 237–238
| Seridji, R. 1988: 1298 |
| Holthuis, L. B. 1956: 238 |
Albunea symnista
| Kensley, B. 1981: 35 |
| Barnard, K. H. 1950: 405 |
Albunea
| Menon, M. K. 1943: 331 |
A. symnista
| Ward, M. 1942: 52 |
Albunea symnista
| Miyake, S. 1978: 152 |
| Gordon, I. 1938: 187 |
Albunea symnista
| Asakura, A. 1995: 376 |
| Miyake, S. 1991: 157 |
| Gordon, I. 1938: 187 |
Albunea symnista
| Miyake, S. 1978: 152 |
| Gordon, I. 1938: 187 |
Albunea symnista
| Gordon, I. 1938: 187 |
Albunea symnista
| Sakai, T. 1935: 61 |
Albunea symnista
| Kikuchi, K. 1932: 10 |
Albunea symnista
| Kikuchi, K. 1932: 10 |
| Nakazawa, K. & I. Kubo & S. Miyake 1951: 741 |
| Nakazawa, K. & I. Kubo & S. Miyake 1949: 741 |
| Nakazawa, K. 1927: 1051 |
Albunea symnista
| Rathbun, M. J. 1924: 29 |
Albunea oxyophthalma: Southwell, 1910: 184
| Southwell, T. 1910: 184 |
Albunea symmysta: Nobili, 1906: 142–143
| Ramadan, M. M. 1936: 3 |
| Nobili, G. 1906: 143 |
Albunea symmysta:
| Holthuis, L. B. 1956: 238 |
| Ortmann, A. E. 1901: 1276 |
| Ortmann, A. E. 1896: 224 |
Albunea symnista
| Cano, G. 1889: 95 |
| Cano, G. 1889: 263 |
Albunea symnista
| Ozorio, B. 1888: 186 |
Albunea
| Claus, C. 1886: 69 |
| Claus, C. 1885: 69 |
Albunaea [sic] symnista
| Stimpson, W. 1858: 230 |
Albunea symnista
| Gibbes, L. R. 1850: 24 |
Albunea symnista
| Gibbes, L. R. 1850: 187 |
Albunea
| Dumeril, C. 1816: 431 |
| Rivera, A. 1933: 22 |
| Rivera, A. 1933: 1 |
| Ferrer Aledo, J. 1914: 68 |
| Bolivar, I. 1892: 128 |
| Barrois, T. 1888: 18 |
| Heller, C. 1863: 153 |
| Lucas, H. 1849: 27 |
| Rafinesque-Schmaltz, C. S. 1814: 20 |
Cancer lymnista
| Froriep, L. 1806: 183 |
Albunea dorsipes: Herbst, 1804: 31
| Herbst, J. F. W. 1804: 31 |
Albunea symnista
| Sakai, K. 1999: 9 |
| Naiyanetr, P. 1998: 51 |
| Calado, T. C. dos 1997: 17 |
| Subramoniam, T. 1993: 133 |
| Seridji, R. 1988: 1298 |
| Calado, T. C. dos 1987: 43 |
| Subramoniam, T. 1984: 78 |
| Naiyanetr, P. 1980: 22 |
| Naiyanetr, P. 1978: 333 |
| Thomassin, B. A. 1969: 138 |
| Sarojini, S. 1962: 191 |
| Boschma, H. 1955: 14 |
| Gurney, R. 1942: 263 |
| Gravely, F. H. 1941: 75 |
| Bouvier, E. - L. 1940: 181 |
| Gordon, I. 1938: 187 |
| Boschma, H. 1937: 204 |
| Menon, M. K. 1937: 10 |
| Boschma, H. 1931: 351 |
| Southwell, T. 1910: 183 |
| Henderson, J. R. 1893: 338 |
| Stebbing, T. R. R. 1893: 152 |
| Ortmann, A. E. 1892: 536 |
| Muller, F. 1890: 472 |
| de Man, J. G. 1887: 425 |
| Albert, F. 1883: 523 |
| Lucas, H. 1881: 54 |
| Miers, E. J. 1878: 326 |
| Heller, C. 1865: 72 |
| Heller, C. 1863: 153 |
| Lucas, H. 1853: 45 |
| White, A. 1847: 57 |
| Milne Edwards, H. 1837: 111 |
| Milne Edwards, H. 1837: 203 |
| Brewster, D. 1832: 234 |
| Latreille, P. A. 1831: 56 |
| Guerin Meneville, F. E. 1829: 12 |
| Desmarest, A. - G. 1825: 173 |
| Desmarest, A. - G. 1823: 283 |
| Lamarck, C. de 1818: 224 |
| Latreille, P. A. 1806: 44 |
| Herbst, J. F. W. 1804: 31 |
| Fabricius, J. C. 1798: 397 |
| Weber, F. 1795: 94 |
Cancer dorsipes: Herbst, 1791: 5–8
| Herbst, J. F. W. 1791: 8 |
Cancer
| de Villers, C. 1789: 157 |
Cancer
| de Villers, C. 1789: 157 |
Hippa symnista
| Fabricius, J. C. 1793: 474 |
| Fabricius, J. C. 1787: 329 |
Hippa symnista
| Fabricius, J. C. 1787: 329 |
Cancer gymnista
| Statius Muller, P. L. 1775: 1128 |
Cancer
| Houttuyn, F. 1769: 422 |
Cancer symnista
| Linnaeus, C. 1767: 1053 |