
Linguamyrmex vladi Barden & Grimaldi, 2017
|
publication ID |
https://doi.org/ 10.1111/syen.12253 |
|
publication LSID |
urn:lsid:zoobank.org:pub:40D636A3-4D88-470A-BC5B-85ABFD1A49E2 |
|
DOI |
https://doi.org/10.5281/zenodo.6004742 |
|
persistent identifier |
https://treatment.plazi.org/id/2C55A44F-974D-4A33-BDC2-26269687577A |
|
taxon LSID |
lsid:zoobank.org:act:2C55A44F-974D-4A33-BDC2-26269687577A |
|
treatment provided by |
Plazi |
|
scientific name |
Linguamyrmex vladi Barden & Grimaldi |
| status |
sp. nov. |
Linguamyrmex vladi Barden & Grimaldi sp.n.
http://zoobank.org/urn:lsid:zoobank.org:act:2C55A44F-974D- 4A33-BDC2-26269687577A
Figs 1 View Fig. 1 , 2 View Fig. 2 A, 3F–I, 5.
Diagnosis. As for genus.
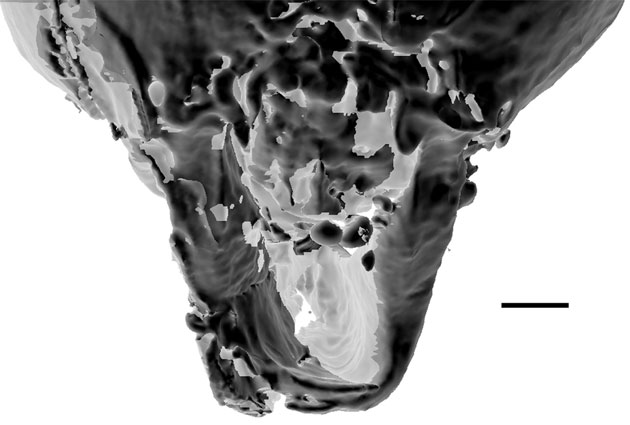
Description. Head: measuring 0.90 mm postero-anteriorly along dorsal margin, 0.96 mm in length/depth from vertex of head to anterior margin of clypeus. Occipital foramen positioned highly dorsad, just under vertex of head. Postgena broadly depressed; postgenal suture visible, deeply furrowed. Vertex broadly rounded and glabrous with gena gradually tapered ventrally towards mandibular socket and oral opening with fine, sparse setae. Ocelli faintly visible on vertex, positioned dorsally. Eye situated high on head capsule and bulging in frontal view, ovoid, measuring 0.38 mm in length and 0.25 mm in width when viewed laterally. Three antennal segments fully preserved (scape 0.94 mm in length; pedicel 0.12 mm; flagellomere I 0.55 mm). Antennal socket approximately in line with ventral margin of eye; socket exposed and immediately flanking a medial frontal triangle (sensu Perrichot et al., 2016). Frontal triangle extends the vertex, contrasted with antennal sockets, which are present within cuticular depressions. Clypeal horn originating at both frontal triangle and clypeal stalk, both structures heavily sclerotized with cleared, membrane-like cuticle connecting from frontal triangle to stalk. Horn paddle-shaped, total length 0.64 mm, diameter 0.49 at greatest; narrow (0.04 mm) glabrous stalk 0.22 mm in length leading to setose pad; setose pad (0.42 in length) with long trigger hairs originating at pad base; dorsal margin of setose pad glabrous, underside coated in a large number of stout setae in centre and longer, more tapered setae along edges. Anterior margin of clypeus medially triangulate; distinct medial ridge extending to clypeal horn; lateral margins, beginning just above mandibular insertion, extending diagonally toward clypeal horn. Cheek-like lobes projecting anteroventrally above mandible insertion. Mandible scythe-like, comprise a linear basal margin (0.55 mm in length) and curved apical tooth (0.92 mm measured as a straight line from base to tip, ignoring curvature) meeting nearly at right angle; preserved with apical teeth in parallel, nearly touching. Basal portion of mandible with anterior flange-like expansion, concave inner margin coated with pointed setae; leading edge of anterior flange expansion smooth; apical tooth rounded broadly with slight point. Maxillary and labial palps not visible.
Mesosoma. Pronotum broad, coated in fine setae; propleuron reduced, not visible in lateral view except where abutting head capsule anteriorly. Pronotal length 0.72 mm, measured along dorsal margin. Mesonotum 0.34 mm; metanotum 0.24 mm; propodeum 0.49 mm. Weber’s length 1.77 mm, mesosomal height 0.56 mm measured perpendicular to Weber’s length line at pronotum. Procoxal length 0.72 mm, max width 0.22; mesocoxal length 0.31, max width 0.21; metacoxal length 0.49, max width 0.26; protrochantal length 0.14, max width 0.11; mesotrochantal length 0.19, max width 0.11; metatrochantal length 0.19, max width 0.12; profemoral length 1.01, max width 0.13; mesofemoral length 1.09, max width 0.12; metafemoral length 1.48, max width 0.18; protibial length 1.09, max width 0.11; mesotibial length 1.30, max width 0.08; metatibial length 1.59, max width 0.08. Trochantellus present, length included in femur measurements. Three protibial spurs present, the largest approximately 2 × length of other two; mesotibia with two spurs, the larger 2 × the length of the smaller; two conspicuous setae of equal length positioned along anterior margin of mesotibial apex; metatibia with two spurs, the largest pectinate and ~ 3 × the size of the smaller. Pretarsal claw with subapical tooth positioned closer to apex than to claw origin. Dorsal margin of propodeum gradually rounded, posterior margin with sheer face. Propodeal spiracle a dorsoventrally elongate slit situated medially. Metapleural gland opening gaping, present within horizontal invagination of cuticle, opening visible posteroventrally following well-developed bulla.
Metasoma. Petiole 0.49 mm in length, 0.49 mm in height at greatest, pedunculate; not tergosternally fused, deep suture visible; anterior margin gradually sloping; with gradually rounded dorsal margin; broadly attached to gastral segment I (abd seg III) with striated helcium clearly visible. Lateral sulcus visible running anteroposteriorly along petiole, signifying incomplete fusion; ventral to sulcus lies thin, membraneous cuticular expansion with distinct anteriorly positioned keel. Sternite III with pointed projection on anterior margin exhibiting striations running laterally. Circumgastral constriction present between gastral segments I and II with dorsal, posteriorly extended notch. Gastral cuticle translucent, revealing heavily sclerotized and expanded apodemes of gastral sternites IV and V withdrawn into the gaster.
Type material. Holotype, AMNH BuPH-01, in the Department of Invertebrate Zoology, American Museum of Natural History.
Additional material examined. Although not named, three specimens, BuPH-02 ( Figs 2 View Fig. 2 D, 3C, 7, Video S1), BuPH-03 ( Figs 2 View Fig. 2 B, 3A, B, 4) and BuPH-04 (2C), were studied and are illustrated here. These three specimens probably represent additional species, particularly with respect to overall size and clypeal paddle composition. Moreover, these specimens possess mandibles that appear to interlock basally, joining the distal apical tooth portions of the mouthparts ( Figs 3 View Fig. 3 D, E, 7, Video S1). As the inner surface of each mandible is concave, their joining forms a channel that is open dorsally near the pointed mandibular apex ( Fig. 7 View Fig. 7 ). However, there remains some ambiguity regarding the reliability of measurements and identification of other discrete features due to incomplete preservation. We therefore delay description until more complete material is obtained. Measurements are provided here (in mm): (BuPH-02) [BuPH-03] {BuPH-04} Head length measured postero-anteriorly along dorsal margin (1.14) [0.87] {0.69}, head depth (1.48) [1.22] {1.2}, scape length (1.56) [1.36] {1.28}, paddle length (1.5) [1.52] {1.26}, paddle diameter (−) [~ 0.57] {−}, eye length (−) [−] {0.29}, eye width (−) [−] {0.19}, basal margin of mandible (0.71) [0.53] {0.39}, apical tooth (1.97) [1.57] {1.48}, pronotal length (1.08) [0.97] {0.8}, mesosomal height (~ 0.81) [~ 0.75] {0.66}, Weber’s length (2.64) [2.49] {1.99}, petiole length (0.82) [0.76] {0.60}. Unfortunately, no specimen retained all antennal segments, preventing a reliable comparison of scape length relative to all other segments. In addition, all specimens, including the type, are missing terminal abdominal segments.
Etymology. In reference to Vlad III, or Vlad Dracula (c. 1429–1476), prince of a region of Romania then called Wallachia. His moniker, Vlad the Impaler, refers to his favoured and frequent method of execution, which inspired the vampirous character Count Dracula fictionalized by Bram Stoker in 1897. The patronym is in reference to the presumed impalement of prey by Linguamyrmex and its liquid diet (see later).
Systematic comments. Haidomyrmecines have been recovered as monophyletic ( Barden & Grimaldi, 2016) and have traditionally been placed as tribe within Sphecomyrminae ( Bolton, 2003) . The subfamily is defined largely by plesiomorphic features and is now suspected to be paraphyletic ( Barden & Grimaldi, 2016). We tentatively follow this subfamilial assignment here but note that, following additional phylogenetic analyses, haidomyrmecines may be best served by the erection of a separate subfamily.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.