
Seira atlantica, Negri, Ilaria, Pellecchia, Marco & Fanciulli, Pietro Paolo, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.159509 |
|
publication LSID |
lsid:zoobank.org:pub:84F61E38-4DFE-4767-B938-C8A2CF7379FC |
|
DOI |
https://doi.org/10.5281/zenodo.6265803 |
|
persistent identifier |
https://treatment.plazi.org/id/9E288781-FFFD-FFB6-1026-D796B544FCB2 |
|
treatment provided by |
Plazi |
|
scientific name |
Seira atlantica |
| status |
sp. nov. |
Seira atlantica sp. n.
Sampling locality
Near Zad Pass, Middle Atlas, 2100 m , Fig. 1 View FIGURE 1 .
Types
Holotype and 16 paratypes have been deposited in the collembolan collection of Prof. R. Dallai at the Department of Evolutionary Biology of the University of Siena, Italy.
Derivatio nominis
From Atlas , the ancient Latin name of the Moroccan mountains.
Description
Blue pigment in the eye patch and along the antennal segments I, II and III. On the head the blue pigment also located in the clypeal, occipital regions and on the legs, from the subcoxae to the femurs but absent from the tibiotarsi. The body is yellowish in alcohol; some individuals show a weak blue pigmentation in the lateral side of the IV abdominal segment and in the ventral manubrium. The body is covered by brown scales which give a darker appearance to the living animals ( Fig. 2 View FIGURE 2 A). The scales also occur on the dorsal surface of the Ant I, all surface of Ant II and all surface of the proximal half of Ant III.
Body 2,7 mm long (measurements and ratios from the holotype). Antennae are about 2 mm long and the single segments are: Ant I=315 µm, Ant II=510 µm, Ant III=495 µm and Ant IV=730 µm (ratio 1:1,63:1,57:2,5); antennae do not show any degree of annulation. Antennal segment IV with a single terminal vesicle ( Fig. 3 View FIGURE 3 F); antennal organ III consists of 2 sensory rods in a cuticle fold ( Fig. 3 View FIGURE 3 G). Cephalic diagonal 810 µm, and the ratio with antennal length is 2,85. Abdominal tergites III and IV are respectively 270 µm and 1080 “m long; their ratio is 4. Manubrium is 765 µm long while the densmucro length reaches 900 µm; apical part of dens and mucro as in Fig. 3 View FIGURE 3 B. Retinaculum with 4 teeth and 1 macrochaeta on the corpus. The length of femur, tibiotarsus and claw of the third leg is 630 “m, 830 “m and 110 “m respectively. Claw with 2 proximal and 2 distal teeth ( Fig. 3 View FIGURE 3 C). Empodial appendage lanceolate with a small basal tooth ( Fig. 3 View FIGURE 3 C). Trochanteral organ as in Fig. 3 View FIGURE 3 E with more than 30 smooth setae. Ocular plate with 8 + 8 pigmented ocelli. Chaetotaxy of the labrum 4,5,5,4 and labral papillae as in Fig 3 View FIGURE 3 H; chaetotaxy of labium as in Fig. 3 View FIGURE 3 D with formula M1M2REL1L2. Ventral tube showing about 17+17 distal ciliated setae ( Fig. 3 View FIGURE 3 A). Ventral manubrial chaetotaxy with 4 anteapical setae ( Fig. 3 View FIGURE 3 I); 3+3 setae bordering the ventral groove on the head ( Fig. 3 View FIGURE 3 K).
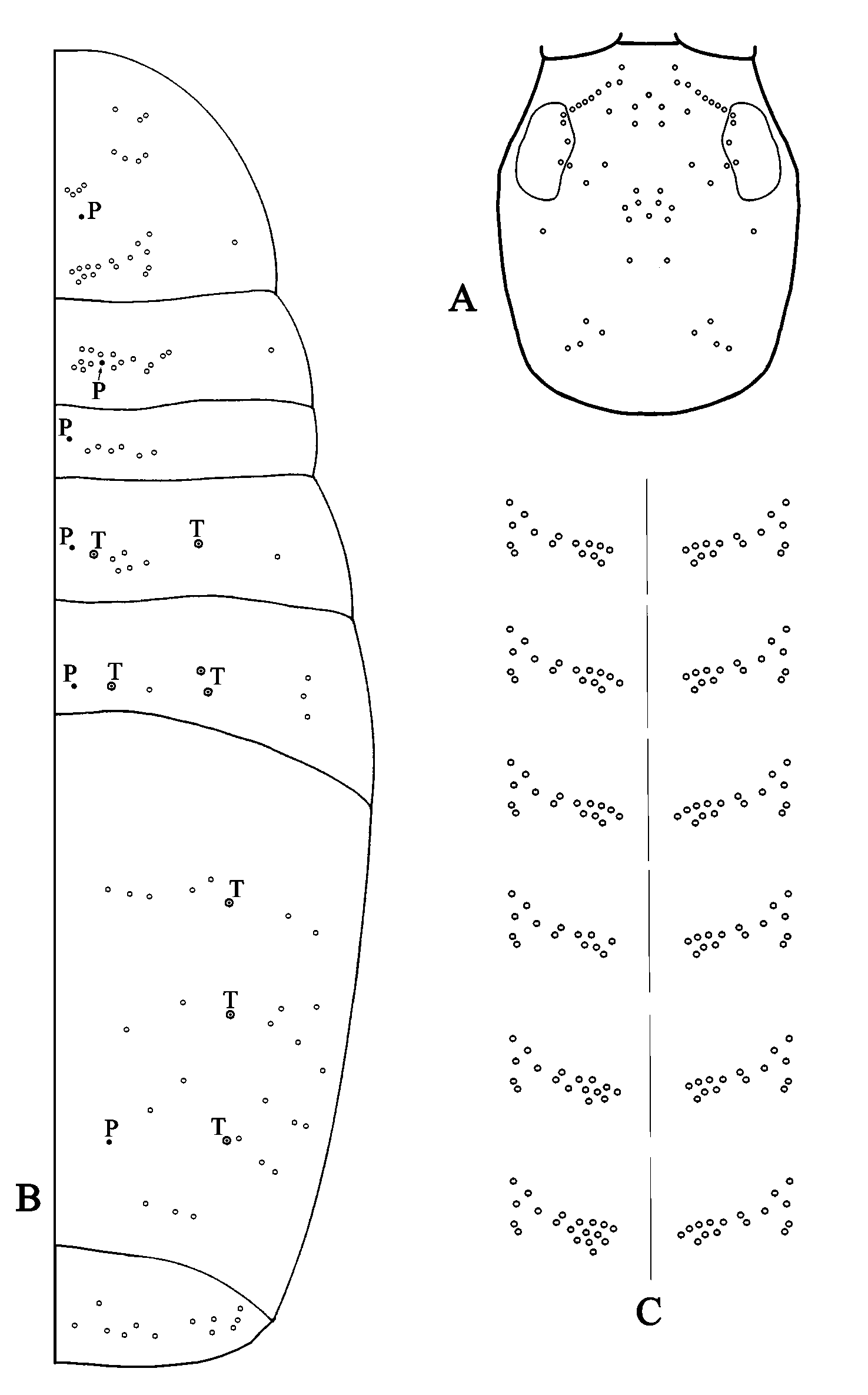
Dorsal chaetotaxy of the head as in Fig. 4 View FIGURE 4 A. The interocular area has 11 multilaterally ciliated macrochaetae. Five macrochaetae are placed in the frontal region. The centralposterior part of the ocular plate (“Zone 3” according to Jacquemart, 1974) shows 4 macrochaetae, while a single one lies inside the ocular plate. In the central area there are 11 macrochaetae. The occipital area shows 4 macrochaetae while there is only 1 macrochaeta posterior to the ocular plate.
Body chaetotaxy as in Fig. 4 View FIGURE 4 B. Thoracic tergite II has 26 macrochaetae on the dorsal region and 1 in the lateral position. Following Jacquemart (1974) the dorsal macrochaetae can be divided in three main zones. “Zone 1” contains two sets of 3 and 4 macrochaetae respectively; “Zone 2” shows 4 “L” forming macrochaetae, and “Zone 3” includes 15 macrochaetae, further distributed as follows: 7 in the “A” group where a certain degree of variability and asymmetry was observed ( Fig. 4 View FIGURE 4 C), 2 in the “B” group, and 6 in the “C” group. Thoracic tergite III has 15 macrochaetae on the dorsal region and 1 in the lateral position; the dorsal ones can be divided into three groups: 7 in group “A”, 4 in group “B” and 4 in group “C”. Tergites of Abd I, II and III with 6(0), 5(1), and 1(3) dorsal macrochaetae respectively (lateral macrochaetae in parentheses). Abd IV shows 12 dorsal macrochaetae distributed according to the following pattern: 5 macrochaetae in the anterior row, 2+ 2 in the central area, and 3 in the posterior row. Abd V with about 12 macrochaetae. Distribution of pseudopores and botriotrichia as in Fig. 4 View FIGURE 4 B.
Discussion
According to Jacquemart (1974), by means of dorsal and cephalic chaetotaxy it is possible to recognise two major subdivisions inside the genus Seira , the S. dollfusi and the S. domestica groups. Moreover, as pointed out by Dallai and Ferrari (1970) and Ellis (1976), within the latter group could be present some synonyms regarding taxa from Mediterranean Basin. On the basis of observed features S. atlantica n. sp. belongs to the domestica species complex. It is easily distinguishable from any other taxa so far described for the simultaneous presence of 7 macrochaetae (with a typical and constant pattern of 4 + 3) in the “A” group of Th III and 5 macrochaetae in Abd II. S. atlantica n. sp. seems to be close to S. dagamae Dallai, 1973 , reported from the Aeolian Archipelago in Sicily. In vivo, the body colour pattern of the two species seems to be quite similar; the cephalic chaetotaxy is the same, as the chaetotaxy of Abd I, III and IV. The number of chaetae in Abd II is also identical (5), but S. dagamae always carries 6 macrochaetae in the “A” group of Th III, instead of 7, which is typical of the new species. However, it should be noted that Ellis (1976) gave the incomplete description of a single female of Seira sp. from Central Crete, whose chaetotactic details are very close to S. atlantica n. sp. We cannot exclude at all that the two taxa belong to the same species, but, as mentioned by Ellis (1976), at present the specimen from Crete cannot be fully evaluated. So, a complete revision of the Seira domestica species complex is strongly recommended.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |