
Parosphromenus juelinae, Shi & Guo & Haryono & Hong & Zhang, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5060.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:6196553C-5180-4E97-8991-99BBC9711AFE |
|
DOI |
https://doi.org/10.5281/zenodo.5608357 |
|
persistent identifier |
https://treatment.plazi.org/id/9D17776C-FA04-FFDF-FF4B-9863FB35F86E |
|
treatment provided by |
Plazi |
|
scientific name |
Parosphromenus juelinae |
| status |
sp. nov. |
Parosphromenus juelinae , new species
( Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
urn:lsid:zoobank.org:act:B3BD985C-28D1-4CFE-B440-9D369055B269
Parosphromenus bintan View in CoL — Kottelat & Ng, 1998: 270 (ZRC30815).
Parosphromenus deissneri View in CoL — Tan & Ng, 2005: 131 (ZRC 46184).
Holotype MZB 25116, male, 19.5 mm SL; Indonesia, Bangka Island , exact locality withheld; colls. Y. H. Ji, 23. Mar. 2017.
Paratypes MZB 25117, female, 16.2 mm SL; NCUMB 65121, 13 specimens, 12.3–19.5 mm SL; same data as for holotype; SJD BA2021–2023, 28 ex. 13.6–32.3 mm SL; same data as for holotype, colls. Y. H. Ji & H. Kishi, 26 Mar. 2017 ; W. T. Shi & J. L. Wang, 30 Sep. 2019 ;
Diagnosis Parosphromenus juelinae , sp. nov., is distinguished from other species of the genus by the following unique combination of characters: the inner iridescent band in the unpaired fins is fragmented into an arc arrangement of several clearly interrupted irregular blotches (when preserved, these iridescent blotches become hyaline); the proximal part of the unpaired fins is black; dorsal-fin rays XIII–XIV, 5–7 (total 18–21, 19*); anal-fin rays X–XII, 9–10 (total 19–22, 21*), caudal fin rounded with a branched media ray, pelvic fin filament light blue; except for the dark brown stripes, the rest of the body is covered throughout by intense red blotches, which extend to the base of the unpaired fins.
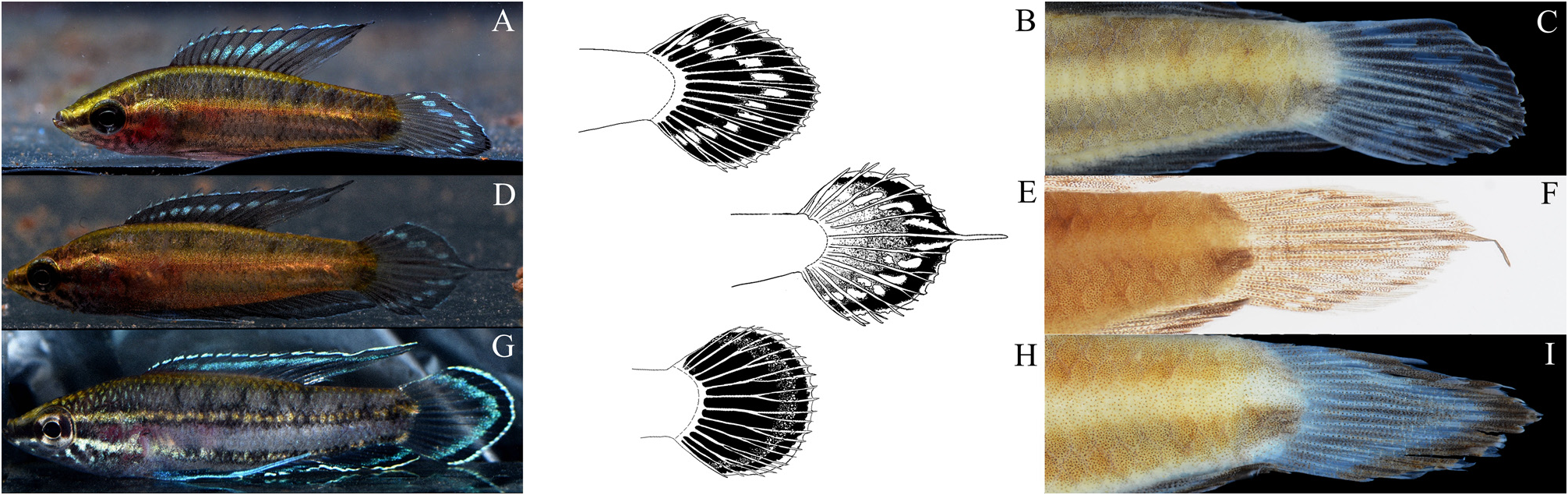
Description Morphometric and meristic data are summarized in Table 1 View TABLE 1 . General body shape and coloration are presented in Fig. 1 A–C View FIGURE 1 . Head pointed, long (25.3%–37.9 % SL); body slightly stocky (22.7%–33.7 % SL, mean 28.1 % at dorsal-fin origin). Dorsal-fin origin usually above 4 th *–5 th scale of lateral scale series. Dorsal-fin with XII 6 (3), XII 7* (8), XIII 7 (2), XIV 7 (1) rays, total 18 (3), 19 * (8), 20 (2) or 21 (1). Dorsal-fin posterior portion pointed distally, reaching beyond caudal-fin base in male, rounded and shorter in female. Caudal fin rounded, with 3 simple, 5 + 6 branched and 2 simple rays. Anal-fin origin below 4 th spine of dorsal fin, posterior portion slightly pointed in male, rounded in female, with X, 9 (1) X, 10 (2), XI, 9 (2) or XI, 10* (9), XII, 10 (1) total 19 (1), 20 (4), 21* (9) 22 (1) rays. Pectoral fin rounded, with 12 (2), 13 (5) or 14 * (8) rays. Pelvic fin with one spine, 1 simple and 4 branched rays, with a long filament reaching about 12 th –13 th anal-fin ray; lateral scales 29 (6), or 30* (9), plus 2 to 3 scales on caudal-fin base; 9 (8) or 10* (7) scales in transverse series upward from 4 th anal-fin spine; 11 (8) or 12* (7) transverse scales at dorsal fin origin.
Live coloration Male ( Fig. 1 B, C View FIGURE 1 ): Head with a yellowish to light brownish background. Dark brown stripes running through flank from snout to caudal peduncle. Sub-orbital and opercular area fully covered by black blotches (less distinct in stressed or preserved specimens). Except for dark brown stripes, body is almost entirely covered by intense red blotches (less intense at anterior part between dorsal-fin base and first dark brown stripe and belly regions). These red blotches turn into a rose-pink tint when fish is stressed; unpaired fins with a bright bluish margin and a narrow black subdistal band; between subdistal band and proximal part, a row of bright turquoise/greenish blotches present, distinctly interrupted by rays of fins ( Fig. 2 A–C View FIGURE 2 ). Proximal parts of unpaired fin mainly black, but red blotches on flank extend to posterior part of dorsal-, anal-fin base and around caudal-fin base. Pelvic fin iridescent bluish, with a bright bluish filament. Pectoral fin hyaline.
Female: Head and body coloration similar to male, but red blotches less intense. Dorsal, anal and caudal fin hyaline without iridescent band. In breeding condition, faint reddish color can be observed in proximal parts of unpaired fins. Pelvic fin filament bluish. Pectoral fin hyaline.
Preserved coloration Male ( Fig. 1. A View FIGURE 1 ): head and body with a whitish or light yellowish ground color (red blotches on body flank in live are not apparent when preserved); a dark brown stripe running from snout through eye and along whole dorsum to caudal-fin base; a second parallel stripe present from postorbital area through flank to middle of caudal peduncle; some black pigments can be observed on suborbital, opercular, and a third short stripe present along belly region to middle part of ana-fin base (up to 8 th spine of anal fin in holotype). Dorsal, anal and caudal fins with hyaline margin; remaining areas of unpaired fins uniformly brownish with a row of hyaline blotches in the middle. Pectoral fin hyaline. Pelvic fin dark brownish with a hyaline edge.
Female: Head and body coloration similar to male but less intense. Unpaired fins light brownish without distinct patterns, hyaline margin present. Pectoral fin hyaline. Pelvic fin base slightly brownish, filament hyaline.
Comparison Parosphromenus juelinae sp. nov. differs from its most similar congener, P. deissneri , in the following combination of characteristics: caudal fin rounded in P. juelinae (vs. lanceolate with a filamentous median ray in P. deissneri ), median ray branched (vs. simple filamentous) ( Fig. 2 D–F; S View FIGURE 2 1 View FIGURE 1 ), base color of the proximal part of the unpaired fins black (vs. dark reddish to brownish), presence of a bluish margin in the posterior part of the median ray of the caudal fin (vs. absence), fewer anal-fin spines (X–XII, XI*, vs. XII–XIII*), a slightly greater body depth at dorsal-fin origin (22.7–33.7% SL, mean 28.1%SL vs. 26.2–27.2, mean 26.8), a shorter dorsal-fin base (29.5–46.1% SL, mean 37.7%SL vs. 38.4–46.0, mean 42.0), a greater interorbital width (25.8–34.9 % HL, mean 30.4%HL vs. 21.0–26.0, mean 23.5). It is also distinguished from other similar species: P. bintan , P. gunawani and P. nagyi , by the presence of intense red blotches on the flank (vs. absence) and a row of interrupted iridescent bluish blotches on the caudal fin (vs. a continuous iridescent band) ( Fig. 2 G–I; S2 View FIGURE 2 ); further, it is distinguished from P. bintan by a distinct dark brown stripe along the belly region to anal-fin base (vs. absence or not distinct), when preserved. It also differs from P. harveyi and P. nagyi by having a median row of hyaline blotches in the dorsal and anal fins (vs. absence), when preserved; further it is distinguished from P. harveyi by the light blue pelvic fin filament (vs. entirely or partly black) and from P. nagyi by the lack of a black blotch or spot in the pelvic fin base (vs. presence). It differs from P. allani and P. barbarae , which also possess intense red blotches on the flank, in the black background color of the dorsal and caudal fins with a row of entirely bluish blotches (vs. reddish fins without such a bluish pattern), and lacking a dorsal or caudal fin ocellus (vs. presence). Parosphromenus juelinae can be further distinguished from P. paludicola by having fewer spines on the dorsal fin (XII*–XIV, vs. XVII*–XIX); from P. linkei and P. pahuensis by the lack of black blotches in the middle of the lateral stripes (vs. presence) and the black base color of the unpaired fins (vs. orange/reddish); from P. ornaticauda and P. parvulus by having more spines in the anal fin (X–XII, XI*, vs. VII–IX) and more spines in the dorsal fin (XII*–XIV, vs. IX–XI); from P. filamentosus by a rounded caudal fin (vs. lanceolate, with a non-branched median filamentous ray) and a black proximal part of the unpaired fins (vs. reddish); from P. sumatranus by the lack of an ocellus on the dorsal fin (vs. present) and a black caudal fin with a row of bluish blotches (vs. uniformly reddish); from P. anjunganensis by a narrow dark subdistal band margined interiorly by a row of bluish blotches in unpaired fins (vs. uniformly reddish); from P. quindecim , by fewer anal-fin spines (X–XI*, vs. XIII), and lack of bluish spots in the proximal part of caudal fin (vs. presence); and from P. alfredi , P. opallios , P. phoenicurus , P. rubrimontis , and P. tweediei by a row of interrupted bluish blotches on the unpaired fins (vs. a partly or completely reddish band).
Distribution Parosphromenus juelinae sp. nov. is restricted to a small area on Bangka Island, Indonesia, where it occurs in a few forest peat swamps with little human interference.
Etymology This species is named after Juelin Wang, who collected the specimens along with Wentian Shi, and whose inspiration and assistance made this study a success.
Field notes This species is restricted to a few adjoining forest streams and swamps belonging to the same river system in Bangka, which are not connected with the habitats of P. bintan and P deissneri . The habitats are wellpreserved and relatively undisturbed by human activities. The water bodies are densely vegetated with aquatic macrophytes (mainly Cryptocoryne bankanensis [ Araceae ] and Utricularia sp. [ Lentibulariaceae ]) and shaded by trees and shrubs at the bank; and the water is clear, with high tannin levels, giving a black-tea color ( Fig. 3 View FIGURE 3 ). As these are less disturbed habitats, the population density was once very high. In 2017, over 100 specimens were collected from a water pool (about 20 m ²) within 40 minutes using three hand nets (60cm × 40cm with 4mm mesh size) in the rainy season. However, the edge of the distribution area of this species is now under growing pressure from agricultural activities. The swamp of the above-mentioned collection location in 2017 has been drained and converted into a paddy field in 2019; thus, likely extirpating the population.
All the syntopic fish species recorded from the habitats include: Channa bankanensis (Channidae) , Eirmotus cf. octozona (Cyprinidae) , Paedocypris sp. , Rasbora einthovenii , Rasbora kalochroma , Sundadanio gargula , Trigonopoma gracile and Trigonopoma pauciperforatum (Danionididae) , Nandus nebulosus (Nandidae) , Belontia hasseltii , Betta chloropharynx , Betta edithae , Betta schalleri , Betta simorum , Luciocephalus pulcher , Sphaerichthys osphromenoides (Osphronemidae) , Hemirhamphodon pogonognathus (Zenarchopteridae) . No other Parosphromenus species were recorded.
Conservation status On Bangka Island, natural lowland habitats are seriously affected by mining activities, oil-palm plantations, and human settlement. During the field surveys on this island from 2016 to 2019, we observed dramatic habitat loss and degradation of natural landscapes, and in 2018 we noticed that the edge of the type locality had been disturbed by agricultural activities. Following the IUCN Red List Categories and Criteria (ver. 3.1), we propose Parosphromenus juelinae sp. nov. be listed as Critically Endangered B2ab (iii), based on its very restricted distribution area in a single river system on a single small island (<50 km 2) with less than three known locations; and the fact that this habitat has been facing direct threats from human activities since 2018. Since there are no significant environmental conservation projects in this area, the survival of this species in the near term is clearly imperilled.
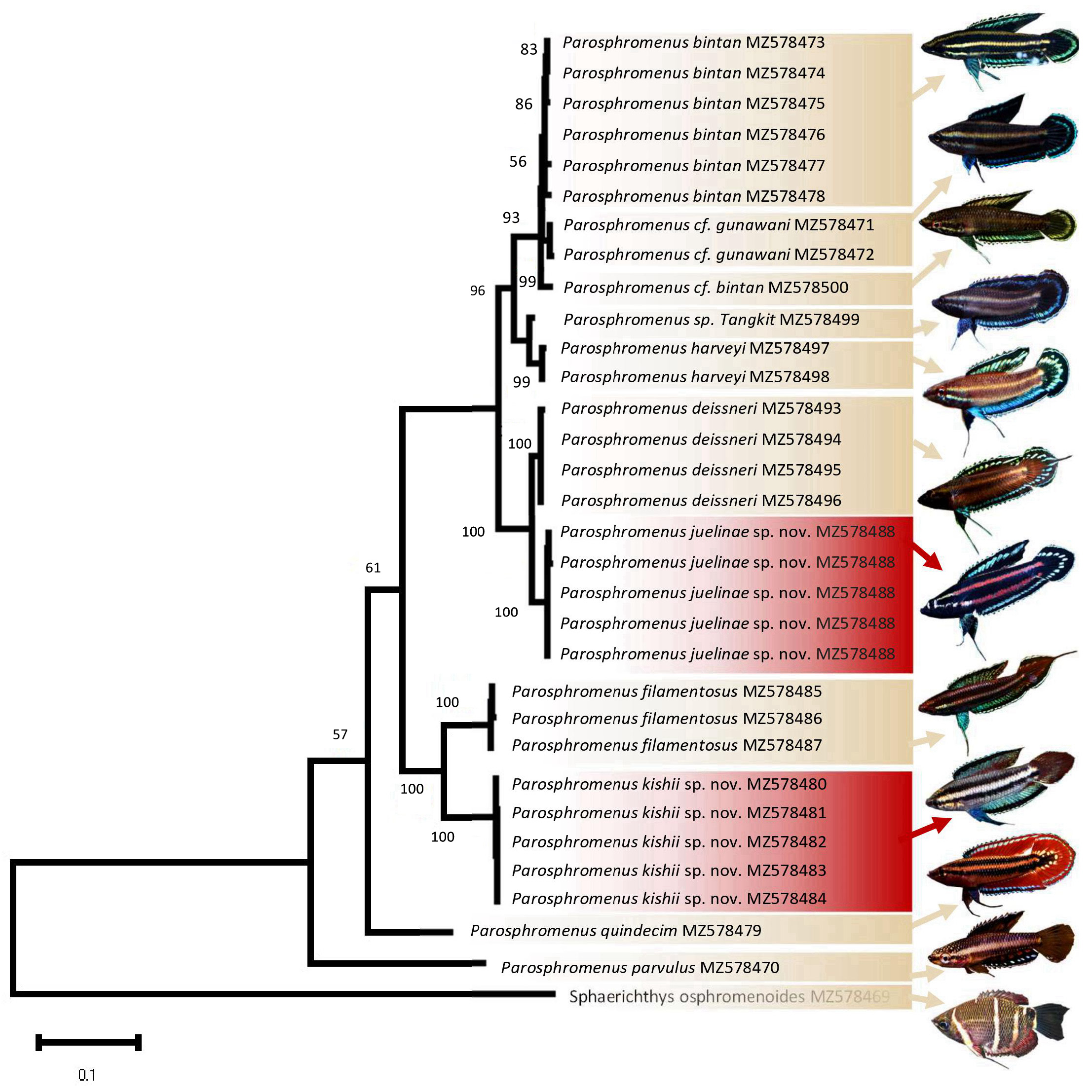
Molecular analysis The consensus phylogenetic tree based on the mitochondrial cytb gene suggests that Parosphromenus juelinae sp. nov. is a monophyletic group distinct from its sister group P. deissneri by an uncorrected p-distance of 2.44% ( Fig. 4 View FIGURE 4 ; Table S2). Parosphromenus . juelinae is significantly distinct from other species, for which cytb data are available with a p-distance ranging from 6.88% to 18.95%. These results indicate that the genetic differences between the new species and its congeners exceed the intraspecific differences observed (<1% in the current study, no published comparable data are known from earlier studies) (K2P <0.5% in related labyrinth fish species like Channa argus, Zhou et al. 2018 ). Morphologically, P. juelinae differs from all known Parosphromenus species (see above Diagnosis and Comparison). Thus, based on both a statistically significant morphological diagnosis based on 43 examples and a cytb divergence consistent with that between other sister-pairs of anabantoids fishes, we are confident that, these specimens from Bangka do in fact represent a valid species.
Remarks Kottelat and Ng (1998) noticed that the specific combination of caudal-fin shape and color pattern to be consistent within a population of Parosphromenus on Bangka Island. Based on this observation, they distinguished P. bintan : the populations with shape1 (caudal fin rounded with branched median ray) and pattern1 (continuous bluish band) from P. deissneri : shape2 (lanceolate with simple filamentous ray) and pattern2 (a row of interrupted bluish oval blotches). There was a controversial single claim by N. Neugebauer and K. Frank in 1994 that P. bintan and P. deissneri are syntopic, viz. different combinations of characteristics could be found in a single population in a location 8 km from Air Bara towards Pajung, which has been cited by Kottelat & Ng (1998) with a note of caution that the specimens mentioned were not examined. This claim was also mentioned by Linke (2014). However, this single report of syntopy in Bangka has not been substantiated by other authors. In 2000, D. Armitage and A. Brown visited this location and recorded only a single species of Parosphromenus ( Armitage 2002) . Further, the Air Bara-Kota Koba area was surveyed regularly by Kishi from 2000 to 2017, but no syntopic of Parosphromenus were found (pers. comm.). In the 2008 survey by H. Linke too, no syntopy was reported around Air Bara ( Linke 2014, pers. comm.). At least since 2017 this river has been severely polluted by Tin mining. We are thus no longer able to substantiate this report. Recently, a local fish conservation organization (Travonim 2021, per. comm.) too, confirmed that only a single species of Parosphromenus could be recorded in the Air Gegas area (Air Bara is a village of this area). Meanwhile, during our own surveys between 2016–2019, we did not find any species of Parosphromenus in syntopy in this region or anywhere else in the island. Our field surveys suggest that each of the species is distributed exclusively in different drainages: P. deissneri in the Sungai Baturusa basin (river system of the original type locality) and Sungai Kurau basin (locality of neotype) towards east coast; P. bintan in the Sungai Kotawaringin basin (our survey 2016) and Sungai Menduk basin (our survey 2017) towards west coast and P. juelinae in rivers draining towards the north coast (location data can be requested from MZB and first author). These results are consistent with Kottelat and Ng’s (1998) observation that characteristics are constant within the population from each specific location.
Thus, there is no reason to discard the principle that the unique combination of shape and pattern of the caudal fin constitutes a diagnostic criterion of a species in this genus. The populations of P. juelinae discovered in distinct drainages can be morphologically distinguished from the other two known species following this criterion based on a different but consistent combination of characteristics: shape1 & pattern2 ( Fig. 2 View FIGURE 2 ). The validity of this morphological difference is further supported by the molecular analysis, which shows that these three species represent three distinct monophyletic groups with characteristic morphological traits.
There are two series of specimens deposited in Lee Kong Chian Natural History Museum, ZRC30815 from Bangka-Belitung and ZRC46184 from ‘Biliton’, Sumatra ( Fig. S3–S View FIGURE 3 4 View FIGURE 4 ) labeled as P. bintan ( Kottelat & Ng 1998) and P. deissneri ( Tan & Ng 2005) , respectively. These are distinct from P. bintan and P. deissneri , but morphologically similar to P. juelinae . A clarification of their taxonomic status is beyond the scope of the current description (See Supplementary Notes of Comparative Materials and Supplementary Figures for more details).


| MZB |
Museum Zoologicum Bogoriense |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parosphromenus juelinae
| Shi, Wentian, Guo, Shujie, Haryono, Haryono, Hong, Yijiang & Zhang, Wanchang 2021 |
Parosphromenus deissneri
| Tan, H. H. & Ng, P. K. L. 2005: 131 |
Parosphromenus bintan
| Kottelat, M. & Ng, P. K. L. 1998: 270 |