
Burmaculex antiquus Borkent & Grimaldi, 2004
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4079.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:3736BD33-404F-442D-B50B-40C19869FCBB |
|
DOI |
https://doi.org/10.5281/zenodo.6089788 |
|
persistent identifier |
https://treatment.plazi.org/id/9C283051-FFB0-FFB2-FF2F-B0C4FB8DD85C |
|
treatment provided by |
Plazi |
|
scientific name |
Burmaculex antiquus Borkent & Grimaldi |
| status |
|
Burmaculex antiquus Borkent & Grimaldi View in CoL
Burmaculex antiquus BorkenT & Grimaldi, 2008: 883 View in CoL . Mid-CreTaceous Burmese amber. Culicidae—Grimaldi, Engel & Nascimbene 2002: 53 (descripTion, illusTraTions, phoTo). Culicidae-Grimaldi & Engel 2005: 506 (discussion, illusTraTions).
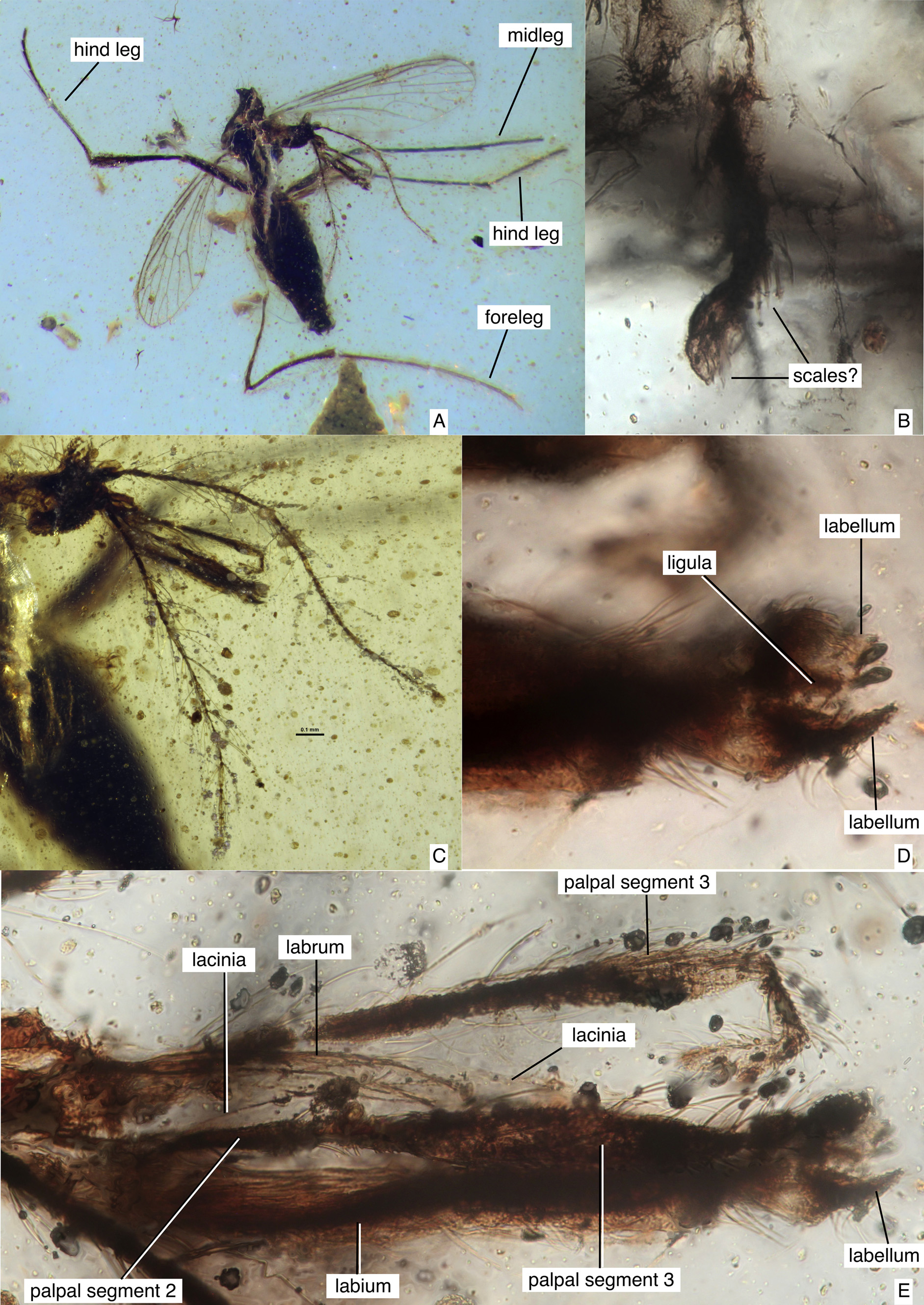
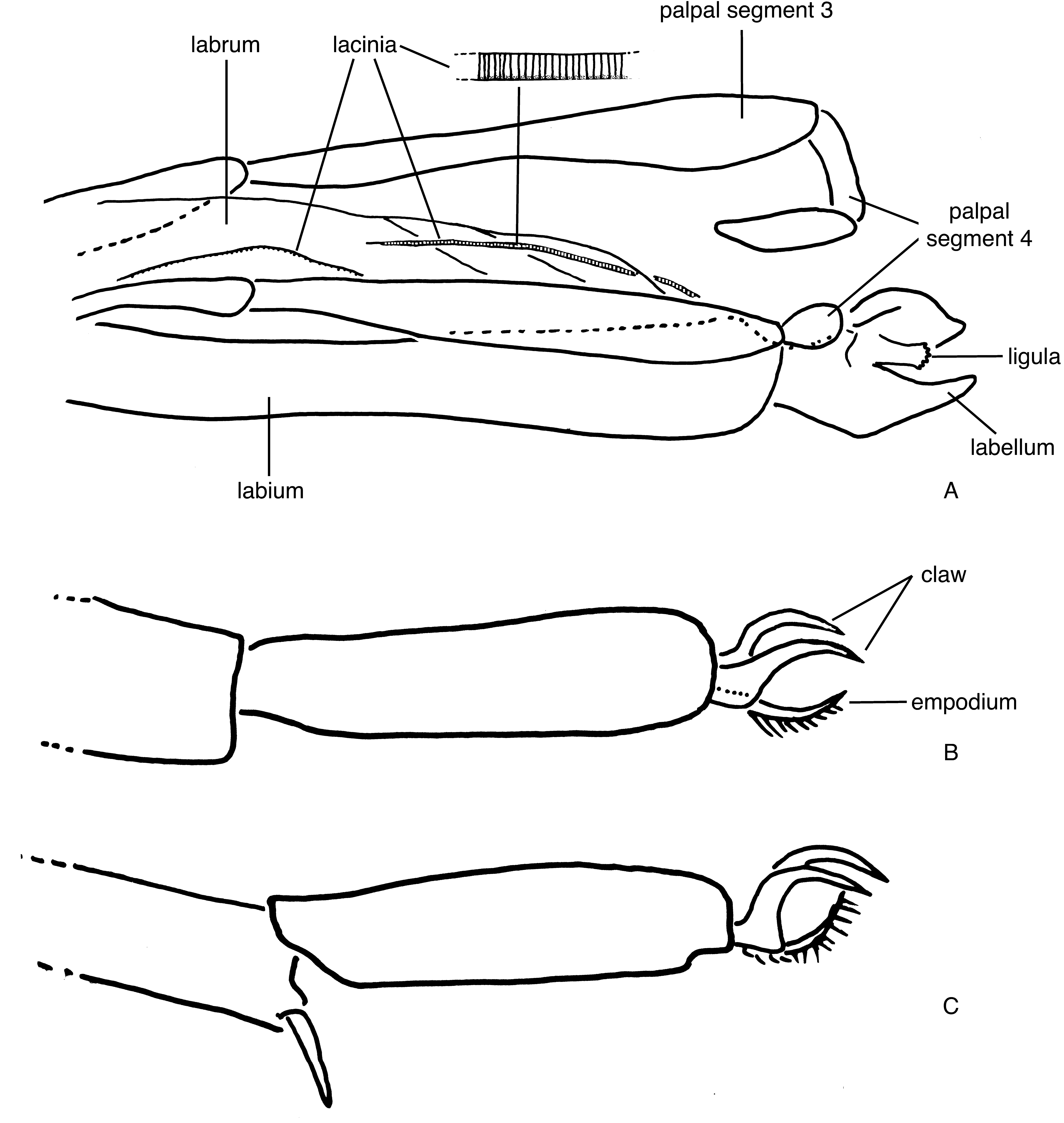
Diagnosis. The only known species of Culicidae with moderately elongate mouthparts (about 0.3–0.4x length of antenna) ( Figs. 1A, C, E View FIGURE 1 , 4A View FIGURE 4 ). Also, the only known culicid lacking scales on the wing veins (scales are present only on the costa and the posterior margin of the wing) ( Figs. 2A–C View FIGURE 2 ), and the only culicid having R1 with an anterior bend anterior to R2+3 ( Fig. 2A View FIGURE 2 ).
The following description includes only those features that differ from or are additional to those by Borkent & Grimaldi (2004).
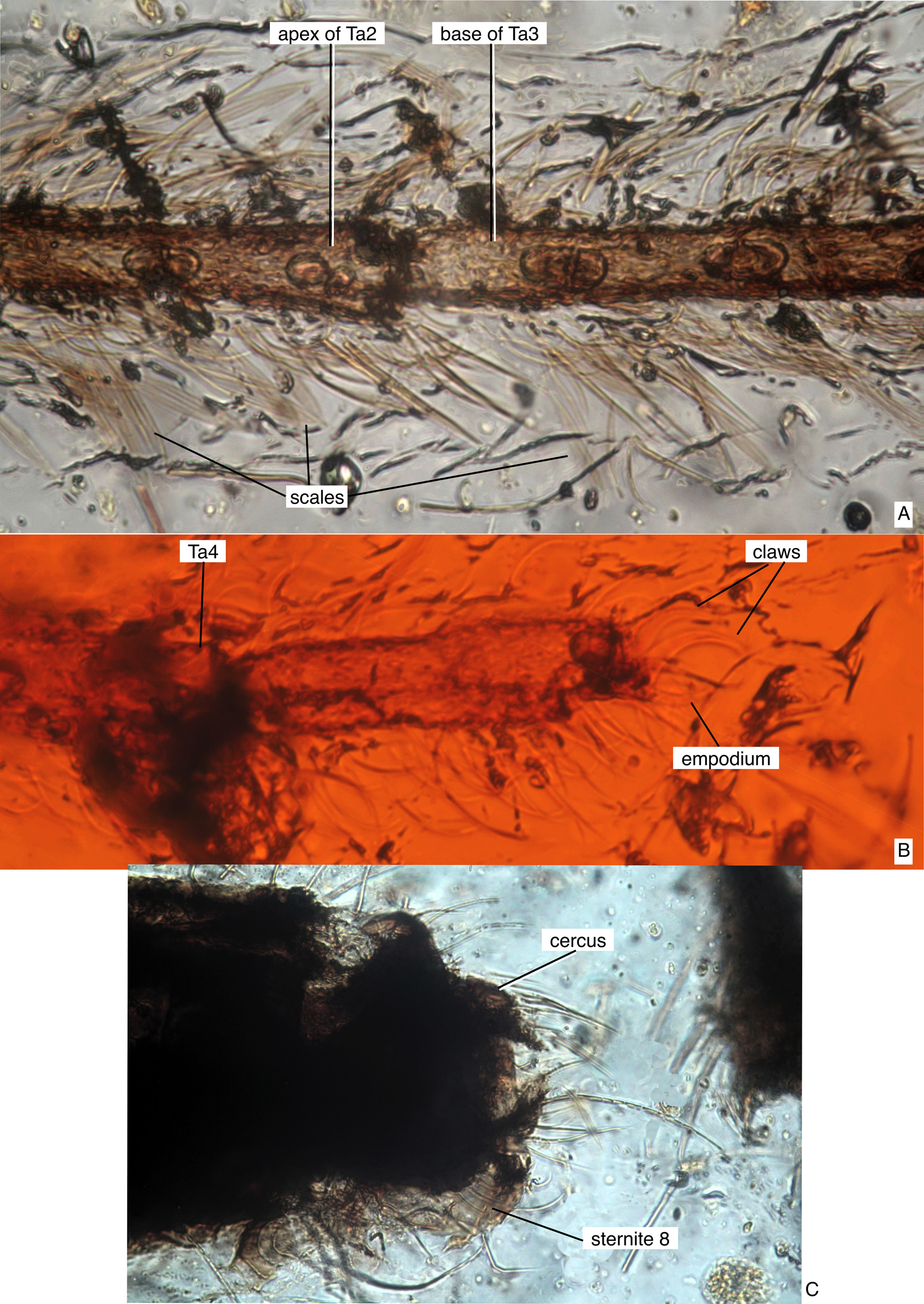
Description (female). Body with scales on legs ( Fig. 3A View FIGURE 3 ), anterior and posterior wing margins ( Figs. 2A–C View FIGURE 2 ), possibly on stem and knob of halter ( Fig. 1B View FIGURE 1 ) (no longitudinal ridges visible on scales). Antenna with flagellomeres 2–12 each with basal whorl of sensilla chaetica arising from ring of pale cuticle ( Fig. 1C View FIGURE 1 ). Mouthparts ( Figs. 1A, C–E View FIGURE 1 , 4A View FIGURE 4 ) moderately elongate (length of labium / length of flagellum = 0.30–0.38). Labrum slender, elongate, curved in cross-section, abruptly tapered apically. Mandible not visible. Laciniae with transverse ridges along length. Labella well-developed, divided medially, details uncertain, ligula with 7 rounded, short apical projections. Wing as in Fig. 2A–C View FIGURE 2 , length = 1.6–1.9 mm, R1 with anterior bend anterior to R2+3, anal vein apex proximal to level of mcu-CuA fork, with slender setae on all veins (but not crossveins), remigium with four elongate setae, scales restricted to anterior (costa), posterior margin of wing, alula, slender microtrichia on membrane. Foreleg with femur, tibia, tarsomeres 1–4 thickly clothed with scales ( Fig. 3A View FIGURE 3 ), Ta1 length 0.96 X Ta2–5 length ( Fig. 1A View FIGURE 1 ); mid-, hind legs with numerous setae, scales; foreleg, hind leg claws without basal tooth ( Figs. 3B View FIGURE 3 , 4B–C View FIGURE 4 ). Fore-, hind legs each with spiculose empodium ( Figs. 3B View FIGURE 3 , 4B–C View FIGURE 4 ). Abdomen with well-developed sternite 8 overlapping base of moderately elongate cerci ( Fig. 3C View FIGURE 3 ).
Taxonomic discussion. This second specimen of B. antiquus is in fairly poor condition ( Fig. 1A View FIGURE 1 ). Unlike the mostly deteriorated and partially cleared first specimen, the fossil at hand is largely opaque, collapsed and distorted. Nearly all of the body is present but there are only three attached legs, the left mid- (missing at least tarsomeres 3-5) and hind legs (one of these missing some terminal tarsomeres), and the right hind leg. A foreleg is near but detached from the body (identified as such by presence of tibial spur).
Some of the mouthparts were visible in the fossil, confirming that the laciniae (both visible) have transverse ridges ( Fig. 4A View FIGURE 4 ) and are slightly thickened mesially, features which are unique among the stylets of extant Culicidae (mandibles lack these features) ( Lee & Craig 1983). The apices of the laciniae and labrum are obscured by the palpi and perhaps are within the apex of the labium. The mandibles are not visible (probably because they are either too close to the laciniae or are yet enclosed by the labium; in extant species they are extremely slender, with apical teeth at most, and are often difficult to locate in slide-mounted specimens).
Borkent & Grimaldi (2004) reported the possibility of a gap between palpal segments 2 and 3 but this is not present in this second specimen, indicating that this is likely an artifact of preservation in the holotype. There is, however, a lightening of the cuticle between segments 2 and 3 of at least the left palpus in this second specimen ( Fig. 1E View FIGURE 1 ). Similarly, a possible gap at the very base of R4+ 5 in the holotype, but not in this second specimen, is also likely an artifact.
The amber piece holding the fossil has been cut into four pieces, with a small flat slab now bearing the specimen. The slab bearing the fossil also includes a trichome and a very small poorly preserved arthropod. The three separated pieces of amber cut away from the fossil include the anterior portion of a myriapod, several trichomes, various distorted parts of arthropods, as well as debris.
The fossil was collected from the same locality as the first B. antiquus , is housed in a small plastic box at the American Museum of Natural History, and is labeled with "BURMESE AMBER (Cretaceous E. Cenomanian), Myanmar: Kachin, James Zigras Collection, JZC-Bu213, PIP: ♀, Culicidae Burmaculex ", " Kachin: Tanai Village (on Ledo Rd.), 105 km NW Myitkyna, Burma " and " Burmaculex antiquus , female, det. Borkent and Grimaldi".
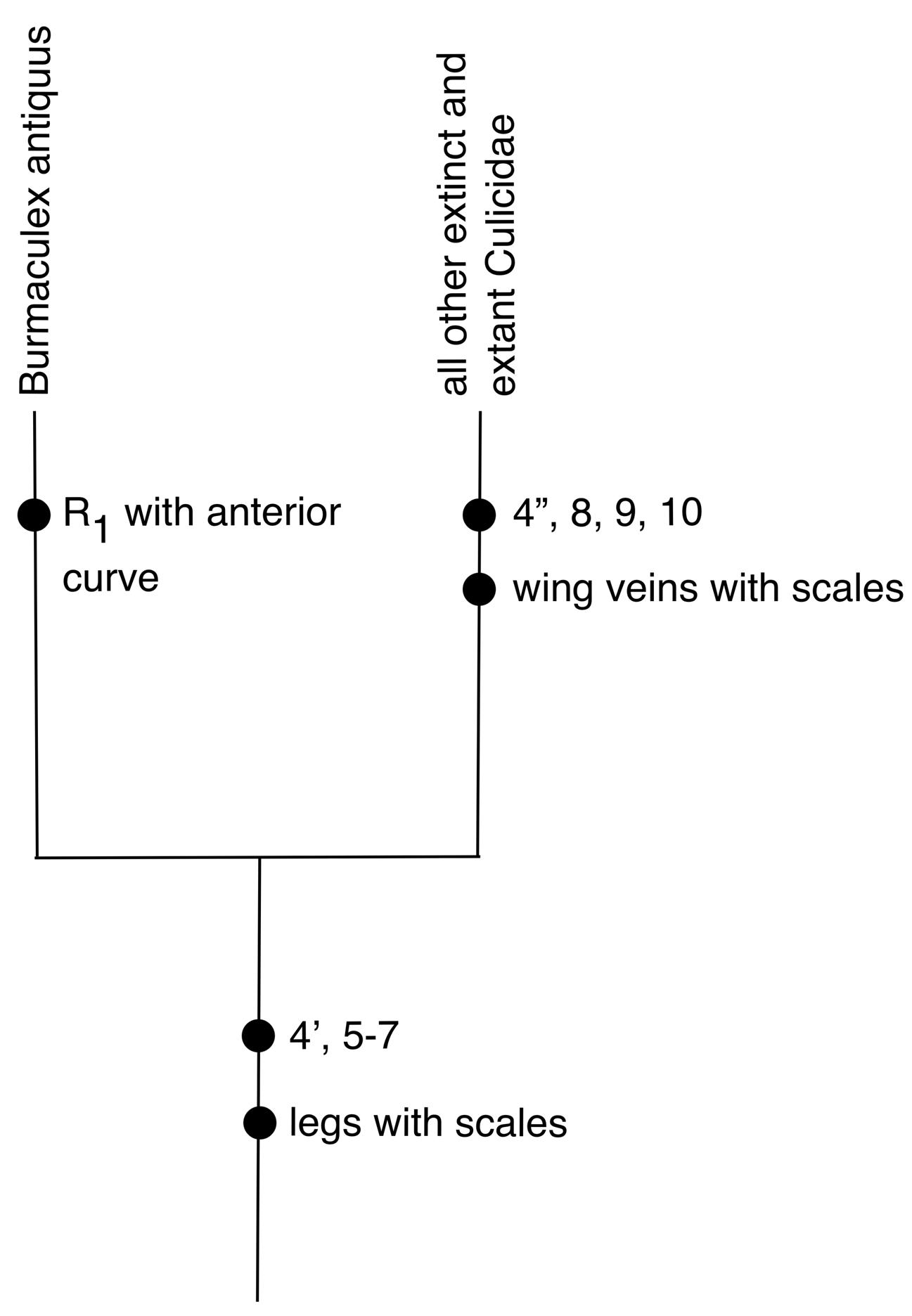
Phylogenetic position of Burmaculex antiquus . Borkent & Grimaldi (2004) provided a phylogenetic analysis of the features of B. antiquus and other Culicidae , with four synapomorphies defining the monophyly of the family including B. antiquus and four synapomorphies defining Culicidae exclusive of B. antiquus . Here we report additional synapomorphies and an autapomorphy of B. antiquus .
The new Burmaculex fossil has abundant scales on the legs, from the base of the femur to at least tarsomere 4 of each leg, and these were particularly evident on the well-preserved foreleg ( Fig. 3A View FIGURE 3 ). This apomorphic state is shared with all other extant and fossil Culicidae ( Fig. 5 View FIGURE 5 ) and is unique within at least the Culicomorpha. Among other nematocerous Diptera , leg scales are also present in some Cecidomyiidae and Psychodidae where they are clearly independently derived.
The presence of scales on the palpus of all Culicidae other than B. antiquus remains a valid synapomorphy (synapomorphy 10 of Borkent & Grimaldi 2004) ( Fig. 5 View FIGURE 5 ). Burmaculex antiquus also lacks scales on the head capsule, thorax and abdomen. Although the latter may be through loss by rubbing and from flowing resin, we consider this unlikely since these surfaces on the insect show no traces of scales while they are abundantly present on the legs. Scales appear to be present on the head capsule and thorax of at least all extant Culicidae (their presence or absence uncertain in some fossils). Most species of Anophelinae lack scales on the abdomen (tergites and sternites usually without scales, sternites always largely devoid of scales) and may have only narrow thoracic scales restricted to the scutellum of the thorax. As such, scales on the head capsule and at least the scutellum may be another synapomorphy of Culicidae other than B. antiquus , but perhaps directly related to the presence of scales on the palpus and wing veins (i.e. they may all be one character–"more broadly distributed scales"). The halter may have scales ( Fig. 1B View FIGURE 1 ) but visibility was limited and this is uncertain. There is likely more phylogenetic information in the specific distribution of body scales in Culicidae but that is beyond the scope of the present paper.
Borkent & Grimaldi (2004) pointed out that the presence or absence of wing vein scales (other than on the wing margin) was difficult to interpret cladistically because scales are present on the wing veins of Chaoboridae and Corethrellidae ( Chaoboridae is the sister group of Culicidae and Corethrellidae of Chaoboridae + Culicidae (Borkent 2012)) . Dixidae lack wing scales. Further information is now available regarding this character. The worldwide revision of Corethrellidae by Borkent (2008) showed that all the early lineages of Corethrellidae have only quite slender scales on their wing veins. Unfortunately, the single specimen of the oldest fossil, from Lebanese amber, Corethrella cretacea Szadziewski , is missing its wings. The earliest extant lineage of Chaoboridae , Eucorethra underwoodi Underwood , has only slender scales on its wing veins. Species of Mochlonyx Loew and Chaoborus Lichtenstein have very narrow scales that could also be called somewhat widened setae. As such, the broad wing scales of all extant and fossil Culicidae other than B. antiquus , is unique and therefore considered a synapomorphy of that lineage. The simple setae on the wing veins of B. antiquus might be considered derived but the simple setae of Dixidae and the rather small difference between the condition in B. antiquus , Mochlonyx , Chaoborus , and early lineages of Corethrellidae suggest this is at best a weak indicator of relationship.
The anterior curve of R 1 in the portion anterior to R2+3 of the wing of B. antiquus ( Fig. 2A View FIGURE 2 ) is unique within Culicomorpha (including all fossils) and is therefore considered an autapomorphy of the species. One of the two wings of the Middle to Late Jurassic Chaoboridae fossil Hypsocorethra toficola Kalugina illustrated by Kalugina & Kovalev (1985: fig. 34b) shows an anteriorly curved R1, but the other wing (their fig. 34a) is shown with this vein straight. The adult mouthparts of this species are relatively short and if the immatures are correctly associated, H. toficola is clearly a member of the Chaoboridae (having larval synapomorphies with other members of the family). As such we consider any potential similarity between the R1 of H. toticola and B. antiquus to be, at best, due to homoplasy.
These hypotheses of character polarity are summarized in figure 5, with character states 4–10 referring to those discussed by Borkent & Grimaldi (2004) and listed as follows:
4. Short proboscis (plesiomorphic); moderately long proboscis ( Figs. 1A, C, E View FIGURE 1 , 4A View FIGURE 4 ) (apomorphic'); very elongate proboscis (apomorphic"). 5. Female labrum relatively short, broad, and dorsal to remaining mouthparts (plesiomorphic); labrum stylate, partially enclosed by the labium ( Figs. 1E View FIGURE 1 , 4A View FIGURE 4 ) (apomorphic). 6. Female lacinia either reduced or as a flattened blade with retrorse hooks (plesiomorphic); lacinia a long, slender stylet bearing fine, flattened, transverse ridges ( Fig. 4A View FIGURE 4 ) (apomorphic). 7. Palpal segment 3 short or of moderate length (plesiomorphic); palpal segment 3 disproportionately elongate ( Figs. 1C, E View FIGURE 1 , 4A View FIGURE 4 ) (apomorphic). 8. Female palpus longer than other mouthpart elements ( Figs. 1C, E View FIGURE 1 , 4A View FIGURE 4 ) (plesiomorphic); palpus equal in length or shorter than other mouthpart elements (apomorphic). 9. Clypeus with setae (plesiomorphic); clypeus without setae (apomorphic). 10. Palpus without scales (plesiomorphic); with scales (apomorphic).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Burmaculex antiquus Borkent & Grimaldi
| Borkent, Art & Grimaldi, David A. 2016 |
Burmaculex antiquus
| Engel 2005: 506 |
| Engel 2002: 53 |