
Muscardinus avellanarius (Linnaeus, 1758)
|
publication ID |
https://doi.org/ 10.5281/zenodo.6604339 |
|
DOI |
https://doi.org/10.5281/zenodo.6604282 |
|
persistent identifier |
https://treatment.plazi.org/id/9B215C43-FFD4-DD13-C965-FD79F59CF63C |
|
treatment provided by |
Carolina |
|
scientific name |
Muscardinus avellanarius |
| status |
|
18. View On
Hazel Dormouse
Muscardinus avellanarius View in CoL
French: Loir muscardin / German: Haselmaus / Spanish: Muscardino
Other common names: Common Dormouse
Taxonomy. Mus avellanarius Linnaeus, 1758 ,
Sweden.
An mtDNA phylogeographic study by A. Mouton and colleagues in 2012 uncovered two primary genetic lineages in M. avellanarius : one lineage occurs in Western Europe and the other in Central and Northern Europe, on the Balkan Peninsula, and in Turkey. The two lineages were estimated to have diverged c.7-7 million years ago during the late Miocene, a period of European faunal turnover discussed under family Gliridae above, and are estimated to have cytochrome-b gene sequence divergence value of 7-7%. According to R. D. Bradley and R. J. Baker in 2001, this value is sufficient to recognize the two lineages as separate species. As noted by Mouton and colleagues, however, additional data from independent gene loci are needed before a decision can be made regarding species-level status, as reliable inferences about population history should be based upon multiple independent loci according to J. C. Avise and R. M. Ball in 1990. Results of Mouton and colleagues also showed that the two primary lineages contained sublineages and there may be at least five historically isolated mitochondrial lineages within what is now recognized as one species, M. avellanarius . Five subspecies of M. avellanarius were historically recognized byJ. R. Ellerman and T. C. S. Morrison-Scott in 1951 and G. B. Corbet in 1978, but there is no congruence between previously recognized subspecies and the five isolated genetic lineages uncovered by Mouton and colleagues in 2012. Monotypic.
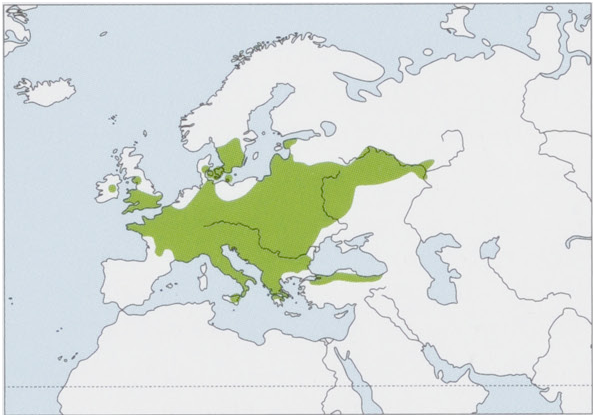
Distribution. W, C, & E Europe, from the British Is (Wales and S England) and France E through most of Europe to the confluence of the Volga and Kama rivers (Russia), NE to Denmark and S Sweden, S to Italy (including Sicily), and SE to N Anatolia (Turkey); a breeding population has been recently confirmed in Ireland. View Figure
Descriptive notes. Head-body 65-91 mm, tail 57-86 mm, ear 9-14-6 mm, hindfoot 14-2-17-8 mm; weight 17-19 g (adults in summer period), but more than 30 g before hibernation. Dorsal pelage of the Hazel Dormouse varies from gray to sandy yellow to golden; it changes in hue as an individual ages. Tail color generally matches that of dorsal pelage, but underside is paler. Ventral pelageis light yellow ocher or yellowish gray. Throat and chest have white patch that continues down to venter in a narrow strip. Tail is furred and bushy, ¢.90% of head-body length. False tail autotomy is characteristic, and individuals with truncated tails are occasionally observed or collected. Individuals with white tail tips can comprise up to 20% of certain populations. Condylobasal length is 18-1-23-4 mm, zygomatic breadth is 11-4-15 mm, and upper tooth row length is 4-1-5-1 mm. Lower jaw has characteristic hole in angular process. External and cranial measurements for adults after the first hibernation are from former Czechoslovakia and Austria. Karyotype is 2n = 46. Females have four pairs of nipples (1 pectoral + 1 abdominal + 2 inguinal = 8).
Habitat. Mostly deciduous or mixed deciduous-coniferous forests with well-developed understories up to elevations of ¢.2000 m. High diversities of tree and shrub species are important determinants of habitat quality for the Hazel Dormouse. A well-developed interconnected understory ensures safety of nest sites from predators, and diversity of understory and overstory species guarantees continuity of food supply in the vicinity of nest sites. The Hazel Dormouse prefers early successional stages of woody vegetation, found in regenerating clear-cut and coppiced areas. Hedgerows can also provide good habitat and serve as corridors between wooded areas. In mountainous areas, Hazel Dormice can live in subalpine dwarf mountain pine ( Pinus mugo, Pinaceae ) forests.
Food and Feeding. Hazel Dormice feed predominantly on vegetation but also on foods of animal origin. Main vegetative foods are buds and flowers in spring, berries in summer, and nuts and soft fruits in autumn. Generative parts of plants are preferred, whereas vegetative parts are only subsidiary foods. During periods of scarcity of suitable vegetable food in late spring and early summer, Hazel Dormice eat food of animal origin, primarily insects, such as aphids and caterpillars, and bird eggs. Insect consumption is expected to be more important in suboptimal habitats characterized by low diversity of nutritious plants.
Breeding. Littersizes of Hazel Dormice are 1-9 young;littersizes of 3-6 are most often recorded, resulting in mean littersize close to 4. Pregnancieslast 22-25 days. Lactation lasts 35-45 days. The Hazel Dormouse is unique among European dormouse species in its capability, even in northern parts ofits distribution, of producing two litters; in exceptional cases, three litters have been recorded in a single season. Young-of-theyear females can breed as early as 2-3 months of age during the season in which they are born. In regions where they hibernate, young are usually born in May-September, but they have been recorded as late as October-November in England. Recent genetic studies indicate that the Hazel Dormouse has a promiscuous mating system where both sexes mate with multiple partners; consequently, young born in the same litter may have different fathers.
Activity patterns. The Hazel Dormouse is nocturnal and crepuscular. Activity commences between sunset and ¢.30 minutes after and ends about one hour before sunrise. Diurnal activity has been recorded during the afternoon, especially in autumn. Daily torporis a characteristic feature of the Hazel Dormouse and occurs most often in spring and autumn. Two patterns of annual biological cycle have been described in Hazel Dormice. The most common pattern exhibited across much of its distribution consists of hibernation period in winter. The second pattern is characterized by absence of hibernation—a pattern exhibited by some Mediterranean populations. Hibernation typically last ¢.6 months in October—April, but duration and timing depend on latitude, sex, and age. Hibernacula are usually located near the surface of the ground, covered only by a thin layer of leaves or moss. During hibernation, body temperatures remain very close to soil temperatures. Hibernation follows a characteristic pattern: torpor bouts up to 30 days are interrupted by bouts of euthermia lasting several hours.
Movements, Home range and Social organization. The Hazel Dormouse is predominantly arboreal and solitary. Adult males and femalesare solitary and have home ranges that partially overlap. Dormice of the same sex, however, usually are antagonistic toward neighbors during the breeding season. Hazel Dormice use c.1 ha annually and exploit different areas within their home range seasonally. Within their home range, individuals use several nests in closed cavities such as tree holes and nest boxes, or in thick tangles of woody vegetation. Hazel Dormice travel typically ¢.150 m (rarely 300 m) per night. Dispersing juveniles can enter home ranges of adults, and there is no hostility between adults and independent juveniles, especially of different sexes. Average density is only 1-2 adults/ha, but up to 5-10 adults/ha in the most favorable habitats. Although the Hazel Dormouse is arboreal, dispersingjuveniles and sedentary adults can move across open areas, moving on the ground for up to 200-500 m and even crossing wide roads. Ultrasonic vocalizations are largely used for social communication by Hazel Dormice.
Status and Conservation. Classified as Least Concern on The IUCN Red List. Nevertheless, the Hazel Dormouse is considered threatened in many regions and is included in Annex IV of the Habitat & Species Directive of the European Union and Appendix II of the Bern Convention. The Hazel Dormouse is included on national Red Lists in many European countries. Habitat loss due to deforestation, woodland fragmentation, and unfavorable forest management are major threats. Particular attention to conservation of the Hazel Dormouse is given in Great Britain, where it is included in the Species Recovery Programme and has been subject to the National Dormouse Monitoring Programme since the 1990s. Large efforts are now being made to protect populations of Hazel Dormice in Germany, Denmark, the Netherlands, and Flanders. Mouton and colleagues in 2012 advocated thatthe five isolated mitochondrial lineages be regarded as independent evolutionarily significant units for conservation management purposes, as outlined by C. Moritz in 1994 and 1999.
Bibliography. Airapetyants (1983), Amori, Hutterer, Krystufek, Yigit, Mitsain, Meinig & Juskaitis (2008), Ancillotto, Sozio & Mortelliti (2015), Ancillotto, Sozio, Mortelliti & Russo (2014), Andera (1987), Avise & Ball (1990), Berg & Berg (1999), Bradley & Baker (2001), Bright & Morris (1996), Bright, Morris & Mitchell-Jones (2006), Bright, Morris & Wiles (1996), Bichner (2008), Chanin & Gubert (2012), Corbet (1978), Eden (2009), Ellerman & Morrison-Scott (1951), Juskaitis (1999, 2005, 2014), Juskaitis & Baltrunaite (2013a), Juskaitis & Blchner (2013), Juskaitis et al. (2013), Kelm et al. (2015), Krystufek & Vohralik (2005), Lozan (1970), Marnell et al. (2013), Mills (2012), Moritz (1994, 1999), Morris (2011), Mortelliti, Amori et al. (2011), Mouton, Grill, Sara, KryStufek, Randi, Amori, Juskaitis, Aloise, Mortelliti, Panchetti & Michaux (2012), Mouton, Grill, Sara, KryStufek, Randi, Amor, Juskaitis, Aloise, Mortelliti, Verbeylen et al. (2012), Naim et al. (2011), Panchetti et al. (2004), Pretzlaff & Dausmann (2012), Ramakers et al. (2014), Rossolimo et al. (2001), Sara et al. (2001), Schulz et al. (2012), Sekeroglu et al. (2011), Spitzenberger & Bauer (2001d), Storch (1978), Verbeylen (2012), Vilhelmsen (2003), Zima et al. (1994).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.