
Eliomys quercinus (Linnaeus, 1766)
|
publication ID |
https://doi.org/ 10.5281/zenodo.6604339 |
|
DOI |
https://doi.org/10.5281/zenodo.6604304 |
|
persistent identifier |
https://treatment.plazi.org/id/9B215C43-FFD3-DD1B-C9C0-F86DFDCFFA3A |
|
treatment provided by |
Carolina |
|
scientific name |
Eliomys quercinus |
| status |
|
28. View On
European Garden Dormouse
French: Lérot d'Europe / German: Gartenschlafer / Spanish: Liron careto europeo
Other common names: Garden Dormouse
Taxonomy. Mus quercinus Linnaeus, 1766 ,
Germany.
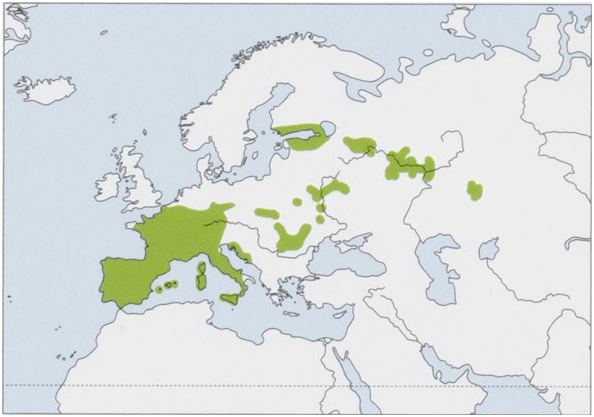
This species historically had a more continuous distribution from Portugal to the Urals in the Russian Federation, but it is presently mostly confined to Western Europe, with eastern populations having become increasingly scattered and fragmented as described by S. Bertolino and colleagues in 2008, M. E. Holden in 2005, and M. G. Filippucci in 1999. Its distribution also encompasses many Mediterranean islands, including Krk, Bra¢, Hvar, Kor¢ula, S¢edro, and Lastovo (off the Dalmatian coast); the Balearic Islands of Formentera, Mallorca, and Menorca; Corsica, Sardinia, Sicily, and Lipari Island. Prehistoric remains, as reported by C. Barbera and M. G. Cimmino in 1990, documented its prior occurrence on Capri Island. As summarized by G. C. L. Perez and colleagues in 2013 and P. Bover and J. A. Alcover in 2008, occurrence of this species on Sardinia and some of the Balearic Islands was actively or passively facilitated by humans. Remains of E. quercinus found in England are likely artifacts from Roman introduction according to D. Yalden in 2010. This species displays a large amount of karyotypic variation across its distribution, and chromosomal number is 2n = 48-54 as summarized by Perez and colleagues in 2013, R. Libois and colleagues in 2012, and E. Gornung and colleagues in 2010. Morphology also varies considerably geographically, as documented by Filippucci in 1999, Filippucci and colleagues in 1988, and B. Krystufek and R. Kraft in 1997. Based on mtDNA analysis, Perez and colleagues in 2013 found that previously described western Palearctic chromosomal races were partially congruent with four primary mtDNA lineages or evolutionarily significant units (ESUs): Iberian, 2n = 48; Italian, 2n = 48 or 50; western European, 2n = 48 or 50; and Alpine, 2n = 52 or 54. Central and eastern European populations have a chromosomal number of 2n = 48. Gene flow persists among chromosomal races; ESUs can be identified by karyotype and mtDNA sequence data but not by morphology and the validity of subspecies names and their geographical limits have yet to be determined. Monotypic.
Distribution. S, W & C Europe (Iberian Peninsula, Balearic Is, most of France, including Corsica, S Belgium, extreme S Netherlands, S & C Germany, W Czech Republic, W Austria, Switzerland, and Italy, including Sardinia & Sicily), and fragmented from E Adriatic coast through E Europe (S Poland and N Slovakia, NE Hungary, Romania, Ukraine, Belarus) to E Ural Mts (Russia), and N to S Finland. View Figure
Descriptive notes. Head—body 99-120 mm, tail 84-110 mm, ear 20-23-5 mm, hindfoot 25-29 mm; weight 33-65-2 g. No sexual dimorphism reported. All species of Eliomys have bold eye masks, but the European Garden Dormouse is generally smaller in body size, has shorter tail and ears relative to head-body length, and has broader interorbit and less inflated bullae relative to skull length, although size varies considerably geographically. Dorsal pelage color is rufous brown or grayish brown with rufous hue and no conspicuous darkening toward midline. Ventral pelage is uniformly white or cream, and dorsal pelage is clearly delineated from ventral pelage. Head color generally matches that of dorsal pelage but with intensified hue. Thick, conspicuous black eye mask extends from above and below ear pinnae, broadly encircles eyes, but does not extend to roots of vibrissae; nose and lips are thinly haired and thus appear almost naked and pink. Ears are brown, moderately long, and rounded; white preand post-auricular patches are present. Hindfeet are white and long, ¢.24% of head-body length. Tail is moderately long, ¢.88% of head-body length. Dorsal tail color generally matches that of dorsal pelage for the first one-half or two-thirds and then abruptly transitions to black. White hairs are scattered throughout, and tail is conspicuously tipped with a broad tuft of white fur; ventral tail color is conspicuously paler. Greatest length of skull is 28-9-33-6 mm, zygomatic breadth is 14-6-18-8 mm, and upper tooth row length is 4-4-5-2 mm. External and cranial measurements listed are of specimens from Slovakia. Females have four pairs of nipples (I pectoral + | abdominal + 2 inguinal = 8).
Habitat. Coniferous, deciduous, and mixed forests and woodland and rocky areas from sea level to elevations of 2300 m in the Alps and Pyrénées. European Garden Dormice also inhabit orchards and gardens and are common in orange groves in eastern Spain. Although most often reported from forest or wooded areas, they are less arboreal than most other dormice and also nest on the ground in rocky areas or karst substrates, scrub vegetation, cracks in stonewalls, and often in houses. In 2007, S. Bertolino and N. Cordero di Montezemolo described the typical Italian Alpine habitat as consisting of abundant rock cover, thick shrub layer, and young understory trees. In 2004, A. Nappi and N. Norante described habitat in the Italian Apennines at 1714-1840 m as being dominated by large cliff-forming rock outcrops and petricolic soils. Habitat selection by European Garden Dormice has mainly been investigated in the Alps. Presence of rocks and stones (useful as refuges from predators and probably for nesting and hibernation), reduced and discontinuous grass cover, and diversified woodylayer positively affect presence and abundance of the European Garden Dormouse in Alpine habitat. They occur in different types of coniferous and mixed forests without availability of rocky cover in the eastern part of their distribution.
Food and Feeding. The European Garden Dormouse is omnivorous. Diet consist of invertebrates, small vertebrates, fruit, nuts, and seeds. Diet analyses reported that they eat insects such as true bugs (Heteroptera), lepidopteran larvae,flies (Diptera), hymenopterans (parasitic wasps, bees, bumblebees, and ants), and beetles (ground beetles, click beetles, weevils, carrion beetles, coleopteran larvae), grasshoppers, and crickets. Other invertebrates recorded are snails (Gastropoda), millipedes and centipedes (Diplopoda), woodlice (Isopoda), spiders ( Araneae ), harvestmen (Opiliones), pseudoscorpions (Pseudoscorpiones), and earthworms ( Lumbricidae ). Prey also includes lizards ( Lacertidae ), the House Mouse ( Mus musculus), the Western Mediterranean Mouse ( Mus spretus), the Long-tailed Field Mouse ( Apodemus sylvaticus), the Greater White-toothed Shrew (Crocidura russula), and songbirds (eggs, chicks, and adults). Plants consumed include fruit of blackberry ( Rubus ssp., Rosaceae ) and elderberry ( Sambucus nigra, Adoxaceae ), hazelnuts, acorns, pine and maple seeds, Phoenician juniper ( Juniperus phoenicea, Cupressaceae ) cones, fruit pulp and peel from oranges, green plant parts, and flowers. Millipedes have odoriferous glands that secrete chlorine, iodine, benzaldehyde, and hydrogen cyanide—a combination that deters most predators—but these and other dormice appear to be resistant to these toxins. There is seasonal variation in the diet.
Breeding. Litter sizes of 1-10 young have been recorded, but litters of 4-6 young are more common. Generally, one litter per year is produced after emergence from hibernation throughout the northern part of the distribution and in montane habitats; two litters per year have been recorded from southern Europe. In eastern Spain, where the climate is mild and food is continually available, European Garden Dormice have been reported to breed throughout the year. Gestation lasts ¢.25 days.
Activity patterns. European Garden Dormice are nocturnal and crepuscular. They enter daily opportunistic torpor to cope with adverse environmental conditions or scarcity of food resources, and throughout much of their distribution, individuals enter periods of hibernation. In areas such as the region of St. Petersburg and in the Alps that experience harsh winters, European Garden Dormice remain in hibernation for as long as 7-5 and seven months respectively, and have resulting activity seasons as short as 4-5-5 months. In regions with milder climates and year-round food availability such as eastern Spain, they are active year-round.
Movements, Home range and Social organization. The European Garden Dormouse is terrestrial and arboreal; it is less arboreal than many other species of dormice. In an alluvial forest, individuals moved primarily on trees and spent one-third of their time on the ground. In montane woodland, individuals reportedly nested and moved mainly on the ground; when they did move through trees, they were 2-6 m above the ground. In the western Italian Alps, densities of 0-2-1-7 ind/ha were recorded during pre-reproductive months and 1-9-4-9 ind/ha after juveniles entered the population.
European Garden Dormice commonly nest in tree cavities and among rocks. They will use bird nests for nesting or food storage; they will either use abandoned nests or consume eggs or nestlings before using an occupied nest; they will also occupy nest boxes. Individuals are thought to use multiple nest sites within a 24hour period. Mean distance traveled between nests was 102 m and did not differ between sexes. The same nest site can be used by 2-3 individuals. Males have larger home ranges than females, and male home ranges overlap with those of females. European Garden Dormice can survive up to five years in captivity but rarely live longer than 2-3 years in the wild.
Status and Conservation. Classified as Near Threatened on The IUCN Red List. Populations of European Garden Dormice have declined significantly in the last 20-30 years; it might have disappeared from as much as 50% ofits former distribution in this timeframe. It might be extirpated in Lithuania and Latvia; it is extremely rare in Estonia, Finland, and neighboring region of St. Petersburg, Russian Federation, because there have been no new records in two or more decades. Populations are likewise declining and in some areas suspected to be extirpated in central and southern Europe. Causes for population decline are not wholly understood. Studies such as that by Perez and colleagues in 2013 that identify evolutionarily significant units are critical for informing conservation measures to preserve genetic diversity and evolutionary potential of the European Garden Dormouse.
Bibliography. Airapetyants (1983), Airapetyants & Fokin (2002), Amori et al. (2016), Andera (1986, 2011), Arnan et al. (2014), Barbera & Cimmino (1990), Bartmanska et al. (2010), Baudoin (1980), Bertolino (2007), Bertolino & Cordero di Montezemolo (2007), Bertolino & Currado (2001), Bertolino, Amori et al. (2008), Bertolino, Cordero di Montezemolo & Currado (2003), Bertolino, Viano & Currado (2001), Bover & Alcover (2008), Crnobrnja-Isailovic et al. (2015), Filipppucci (1999), Filipppucci & Kotsakis (1994), Filippucci, Civitelli & Capanna (1988), Filippucci, Rodin et al. (1988), Gil-Delgado, Cabaret et al. (2006), Gil-Delgado, Mira et al. (2010), Gil-Delgado, Tamarit et al. (2009), Giroud, Turbill & Ruf (2012), Giroud, Zahn et al. (2014), Gornung et al. (2010), Grulich & Jurik (1994), Holden (2005), Juskaitis (1999, 2003), Kahmann & Tiefenbacher (1970), Krystufek & Kraft (1997), Kuipers et al. (2012), Lang (2011), Libois et al. (2012), Macdonald & Barrett (1993), Meinig & Biichner (2012), Moreno (1988, 2002), Moreno & Rouco (2013), Nappi & Norante (2004), Orlov et al. (2013), Perez et al. (2013), Pilats (1994), Rossolimo et al. (2001), Spitzenberger & Bauer (2001b), Storch (1978), Tamarit et al. (2012), Torre et al. (2010), Vaterlaus-Schlegel (1997), Vinals et al. (2012), Yalden (2010), Zima, Macholan, Andera & Cerveny (1997), Zima, Macholan & Filippucci (1994).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.