
Gobiconodon Trofimov, 1978
|
publication ID |
https://doi.org/10.1206/0003-0082(2001)348<0001:GFTECO>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/9A519862-FF9C-FFFB-F661-FD43FBFBFD47 |
|
treatment provided by |
Carolina |
|
scientific name |
Gobiconodon Trofimov, 1978 |
| status |
|
TYPE SPECIES: Gobiconodon borissiaki Trofimov, 1978 .
EMENDED GENERIC DIAGNOSIS: Small to mediumsized triconodonts, with large procumbent i1 and correspondingly enlarged I1; reduction of number of incisors to 1 or 2; conic and pointed posterior incisors, canines, and anterior premolariforms. Anterior premolariforms (p1p3) with tall central cusp and crown height greater than mesiodistal measurement. Accessory cusp on these premolariforms very small to absent. Procumbent ip1. Rounded muscular depression on medial aspect of lower jaw, anteroventral to mandibular foramen (fig. 4; KielanJaworowska and Dashzeveg, 1998: fig. 6c). Separate infraorbital canals connecting orbit with rostrum (see below). Anterior molariform teeth undergo replacement (documented in Gobiconodon ostromi , and probable in the remaining Gobiconodon species ); this feature, however, may be plesiomorphic and not diagnostic. Rounded fossa in the base of the lesser trochanter of the femur and heavier appendicular skeleton (larger crosssection diameter) than other Mesozoic and basal tribosphenic mammals.
Gobiconodon hopsoni , new species
Figures 1 View Fig , 2 View Fig
ETYMOLOGY: After Dr. James A. Hopson, one of a selected few who can comfortably straddle the blurry nonmammalian cynodont/mammalian boundary, with impressive contributions to both segments of therapsid history.
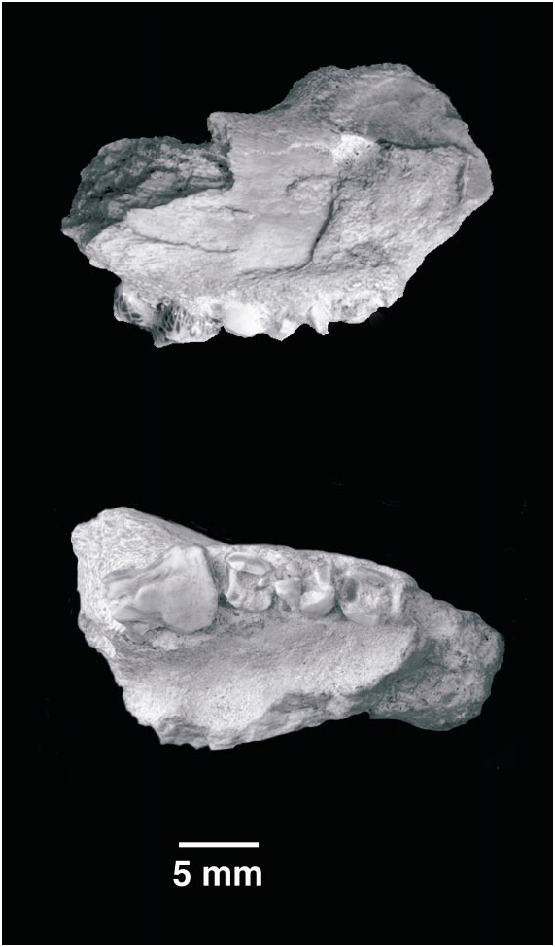
HOLOTYPE: Partial right maxilla ( PSS MAE 140 ) preserving the last molariforms and fragments of the two preceding molariforms.
REFERRED SPECIMEN: PSSMAE 139, a fragment of right lower jaw with a broken, unerupted molariform and alveoli for two more anterior teeth.
TYPE LOCALITY: Oshih, Cannonball Member of Oshih Formation of Berkey and Morris (1927), possibly equivalent of Tevsh Formation (Undurukhin), ValanginianNeocomian (Samiolov et al., 1988). The lower jaw specimens were collected at 44°54'02̎N; 102°53'11̎E.
REPOSITORY: Paleontological and Stratigraphic Section ( PSS) of the Mongolian Academy of Sciences, Ulaan Baatar, Mongolia. The specimens are temporarily stored at the Department of Vertebrate Paleontology, AMNH.
DIAGNOSIS: Large gobiconodontid with deep ectoflexus in a lingually positioned M5. Upper dental arcade forms a closer dental arch than in other species of Gobiconodon , with the last molariforms projecting medially more pronouncedly than in other species of Gogiconodon. Labial cingulum on the M5 broader than that of G. ostromi . Zygoma is deeper in G. hopsoni than in G. ostromi , and projects more posteriorly, closer to the alveolar margin. G. hopsoni is the largest species of Gobiconodon described so far (appendix 1).
DESCRIPTION
MAXILLA (fig.1)
The maxilla is fragmentary, preserving only the root of the zygomatic process, the floor of the orbit, part of the palate, the alveolar margin, and portions of the infraorbital canal.
The lateral aspect of the maxilla is moderately convex without defining a large cheek lateral to the teeth. A small to mediumsized foramen opens laterally at the level of the posterior root of the M3 (fig. 1: Post. Inf.). The foramen leads posteromedially into a canal that opens in the orbit above the orbital platform. We interpret this to be an infraorbital canal, because in mammals the only canal of this size connecting the orbit with the lateral aspect of the rostrum is the infraorbital canal (Sisson and Grossman, 1955; Evans and Christensen, 1979). A portion of another larger canal (fig. 1: Ant. Inf.) can be seen at the broken anterior edge of the maxilla. This canal opens in the floor of the orbital platform as a small infraorbital foramen, lying medial to the infraorbital canal. Judged from the size and orientation of the preserved canal, the more anterior infraorbital foramen was much larger than the more posterior one, and it opened to the rostrum at the level of the M2 or M2M1. Multiple rostral exits of the infraorbital canal are frequent among Mesozoic mammaliaforms (Kermack et al., 1981; Hahn 1985, KielanJaworowska et al., 1987; Lillegraven and Krusat, 1991; Rougier, 1993; Engelmann and Callison, 1998), but we are unaware of other Mesozoic mammaliaforms showing separate infraorbital foramina in the orbit for what are essentially separate infraorbital canals.
The relative size of the infraorbital foramina in this specimen conforms to those of other Mesozoic mammaliaforms, such as morganucodontids (Kermack et al., 1981), basal multituberculates (Hahn, 1985), Vincelestes (Rougier, 1993) , and a Jurassic paurodontid (Hopson et al., 1999), in which the posterior branches of the infraorbital canal are smaller and more ventral than the larger anterior opening.
A faint crest runs along the alveolar edge of the maxilla proximally from the base of the zygomatic process to the broken anterior margin. This crest corresponds to the site of attachment of the buccinator muscle in living mammals (Turnbull, 1970; Evans and Christensen, 1979).
The zygomatic process originates directly lateral to the last molariform. A sizable portion of the process is broken, in particular, the section that would have formed the lower border of the orbit. There is no recognizable facet for the jugal in the preserved portion of the maxilla. A rugose area near the base of the zygomatic process marks the insertion of a strong masseter muscle. The alveolar and zygomatic processes are connected by a gently sloping curve, contrasting with the sharp posterior margin of this area in Gobiconodon ostromi (cf. Jenkins and Schaff, 1988: figs. 4, 5, 7; and fig. 1 here). The pre served morphology of this fragment suggests that the jugal made little contribution to the anterior contour of the orbit and was probably restricted to its posteroventral margin. A substantial part of the anteroventral margin of the orbit was formed by the maxilla, as indicated by the finely rugose, free edge of the maxilla in our specimen. The same condition is present in G. ostromi .
In ventral view, the palatal process of the maxilla exhibits deep palatal fossae for the lower molariforms. As noted also by Kielan– Jaworowska and Dashzeveg (1998), the pattern of the fossae clearly suggests that the occlusion between upper and lower elements alternated; that is, a lower molariform occluded between two upper molariforms. The pattern of the fossae also indicates that a lower molariform occluded behind the last preserved upper molariform. Given that mammalian lower teeth occlude in front of the corresponding upper teeth when the occlusion is alternated, the lower dentition either had one more molariform than the upper dentition, or the small hole observed in the maxilla (labelled as artifact in fig. 1B) represents an alveolus for a more posterior tooth than the one preserved. The latter possibility, however, is unlikely (see comments below).
UPPER DENTITION (fig. 1)
The M5 is the only dental element that is almost completely preserved. A small portion of the distolingual corner of the crown has been chipped away. The specimen is an old individual, and there is heavy wear on the crown. The M5 has two anteroposteriorly compressed roots set widely apart. The crown is subrectangular in occlusal view and was covered by a thick layer of enamel that has been worn down on the lingual aspect of the tooth. Cusp A (fig. 1) is the tallest cusp and is in a central position. Remnants of cusps B and C are anteroposteriorly aligned with A and can be recognized on the crown, but their relative development is uncertain. Cusp C seems to have been slightly more labial in position than either A or B. A moderate, almost unworn labial cingulum extends the length of the labial edge of the crown. The cingulum is quite conspicuous posteriorly, but disappears mesially at about the lev el of the worn cusp B, where it merges into the wear facet. The labial margin of the tooth presents a relatively shallow ectoflexus that divides the margin into two subequal lobes. The lingual side of the crown is either broken or too worn to preserve any evidence of a lingual cingulum. The wear facet, however, slopes mesiolingually from A. The mesial and distal margins of this molariform are slightly concave to flat. The M5 is more lingual than the M4, a condition accentuated in the more distal portion than in the more mesial portion of the M5, resulting in a lingually curving tooth row. The roots of the M4 and M 3 in conjunction with the M5 form a broad, lingually concave arch, suggesting a rapid narrowing of the rostrum in front of the M3 M4 and possibly of the palate behind M5.
The crown of the M4 is missing, but the roots are well preserved. Their transverse section is more circular than the roots of the M5. However, both roots retain a somewhat angular outline. Both M4 and M3 have conspicuous thickenings around the mesial, distal, and lingual aspects of the roots at the alveolar level, which cannot be seen in figure 1. Similar projections of the roots are known in other triconodont lower dentitions, including some amphilestids such as Phascolodon (USNM 2703) , Aploconodon (USNM 2791) , and Gobiconodon borissiaki (PSSMAE collection and Maschenko and Lopatin, 1998: fig. 4c). The M3 is represented only by its distal root and the alveolus of the mesial root.
The slightly triangular outline of the upper molariform crown in gobiconodontids and ‘‘amphilestids’’ (Mills, 1971; KielanJaworowska and Dashzeveg, 1998) was produced by extensive wear along the lingual slopes of cusps A, B, and C. The postnatal ‘‘matching’’ of upper and lower wear facets by extensive remodeling of the crown through wear is a primitive feature of mammaliaforms (Crompton and Jenkins, 1968) retained by Gobiconodon . The extensive wear on the M5 of G. hopsoni , especially of the smaller cusps B and C, imparts to this molariform a somewhat triangular crown pattern.
In ventral view, distomedially from the M5, there is a small circular opening into the substance of the maxilla (Art. in fig. 1). This hole shows no alveolar walls or remnants of teeth. It is also separated from the M5 by a substantial bar of bone, which makes it unlikely that the opening is an alveolus of a more posterior molariform. In addition, the distal aspect of the crown of the M5 has no facet for contact with a tooth occupying a more distal position on the tooth row. It is theoretically possible that the aperture in the maxilla represents a crypt for an unerupted molariform. This interpretation is also unlikely, because of the advanced age of the specimen as shown by the strong wear of the M5. Although late eruption of the last molariforms (Thomas, 1887; Simpson, 1928, 1929; Cifelli et al., 1998) is well documented in triconodonts, the age suggested by the wear in the last preserved molariform seems to be too great to expect subsequent eruption at a new position. Although the opening is in line with the remaining dental elements, we consider it an artifact, and we believe the last preserved molariform to be the last of the series.
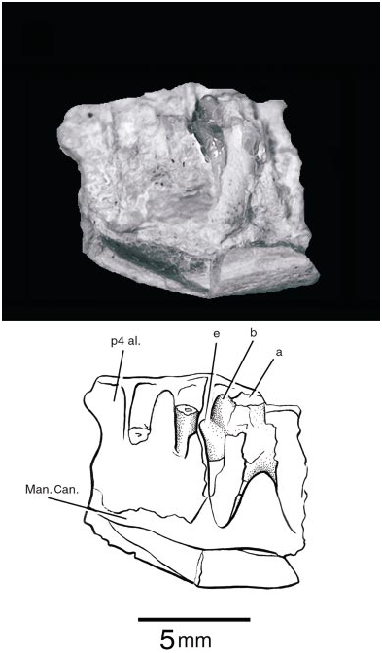
DENTARY (fig. 2)
PSSMAE 139 is a poorly preserved fragment of a right lower jaw found by screen washing. The specimen is also referred to Gobiconodon hopsoni ; the attribution is based on the fact that this specimen is dentally also a triconodont and that it matches in size the maxillary specimen described above. The jaw represents a segment between the anterior and the posterior molariforms, with one partially preserved unerupt ed tooth. This element is likely the m2 and will be so referenced subsequently. In front of the m2 is the partial root of another doublerooted element, and further anteriorly, a larger alveolus for a large singlerooted tooth. The latter alveolus corresponds to the p4 of gobiconodontids, which is variably reduced in G. ostromi (Jenkins and Schaff, 1988) and the gobiconodontids from Khoobur (KielanJaworowska and Dashzeveg, 1998; personal obs.). In G. borissiaki , this tooth position has only one root, but in G. hoburensis the p4 has two roots (KielanJaworowska and Dashzeveg, 1998). On the other hand, G. ostromi (Jenkins and Schaff, 1988) seems to be polymorphic regarding the presence or absence of a p4 because it is present in the specimen MCZ 19860, but absent in MCZ 19965. The p 4 in MCZ 19860 also has contralateral differences; its root is single on one side and ‘‘incipiently double’’ on the other (Jenkins and Schaff, 1988). The differential presence of a p 4 in G. ostromi probably cannot be explained by resorption and plugging of the alveolus by bone as occurs in early mammaliaforms such as Sinoconodon (Crompton and Luo, 1993) , Kuehneotherium (Gill, 1974) , and basal mammals such as Phascolotherium (BMNH 112). The specimen of G. ostromi preserving the alveoli for the p4 (MCZ 19860) appears to be older than the other specimen, which has no evidence of the alveolus (MCZ 19965). In G. hopsoni , the alveolus for the p4 is immediately mesial to that of the m1 and it is not separated from it as in G. ostromi .
The only tooth preserved in PSSMAE 139, the presumed m2, was erupting. In life, the cusp a, the tallest in the crown, would already have cut through the gum. Unfortunately, most of cusp a and the posterior part of the crown are missing. The anterior cingular cusp, e, and cusp b are complete. They are sharp and conical. The lingual aspect of e and most of the anterior slope of b are grooved for the reception of the cuspule d on the distal margin of the preceding molariform. This groove almost reaches the apex of the cusp b, as in Gobiconodon borissiaki but not in G. ostromi . Cusp e is also far more mesial to b than in G. ostromi , but is similar to G. borissiaki in this respect. A cusp f is missing or minimally developed in G. hopsoni . The impression of the distal root shows that it slants posteriorly, as do the distal roots of the posterior molariforms in the Mongolian species of Gobiconodon , particularly in G. hoburensis . G. ostromi , with the exception of the m5, has roots directed more or less vertically. The broken dentary exposes part of the mandibular canal (fig. 2: Man. Can.).
There is a small mental foramen on the labial surface of the dentary at the level between the p4 and m1. The incomplete edge of a second foramen and the small canal leading to it are exposed on the broken anterior margin of the dentary. There are fo fication of the p4 alveolus is correct, then Gobiconodon hopsoni probably had the same pattern of tooth replacement as did G. ostromi , in which the anterior molariforms are replaced. So far, there is no direct evidence supporting a similar pattern in the other known Mongolian species of Gobiconodon (Jenkins and Schaff, 1988; KielanJaworowska and Dashzeveg, 1998) for which several specimens are known, including the modest recent collection by the Mongolian Academy of Sciences and the AMNH. KielanJaworowska and Dashzeveg (1998), however, indicated indirect evidence in favor of molariform replacement among the gobiconodontids from Khoobur.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.