
Waiomys mamasae, Rowe & Achmadi & Esselstyn, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3815.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:F4E00870-86C7-48A5-80B6-871E82767394 |
|
DOI |
https://doi.org/10.5281/zenodo.5116887 |
|
persistent identifier |
https://treatment.plazi.org/id/9A4C8798-FFA4-FFD1-DCEB-F9562AF68ED2 |
|
treatment provided by |
Felipe |
|
scientific name |
Waiomys mamasae |
| status |
sp. nov. |
Waiomys mamasae View in CoL sp. nov.
( Figs. 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Holotype. NMV C37027/ MZB 37000, an adult male collected by hand on the night of 12 May 2012 while it was swimming in a shallow, fast flowing mountain stream ( Figure 2 View FIGURE 2 , 4 View FIGURE 4 ). The specimen was prepared as a dried skin, cleaned skull and fluid-preserved carcass. The carcass, tongue, and phallus were fixed in 4% formalin solution and later transferred to 70% ethanol for permanent storage. The skull was preserved in 70% ethanol, dried, cleaned by dermestid beetles, and degreased by immersion in 10% ammonia. Molars of the specimen are fully emerged.
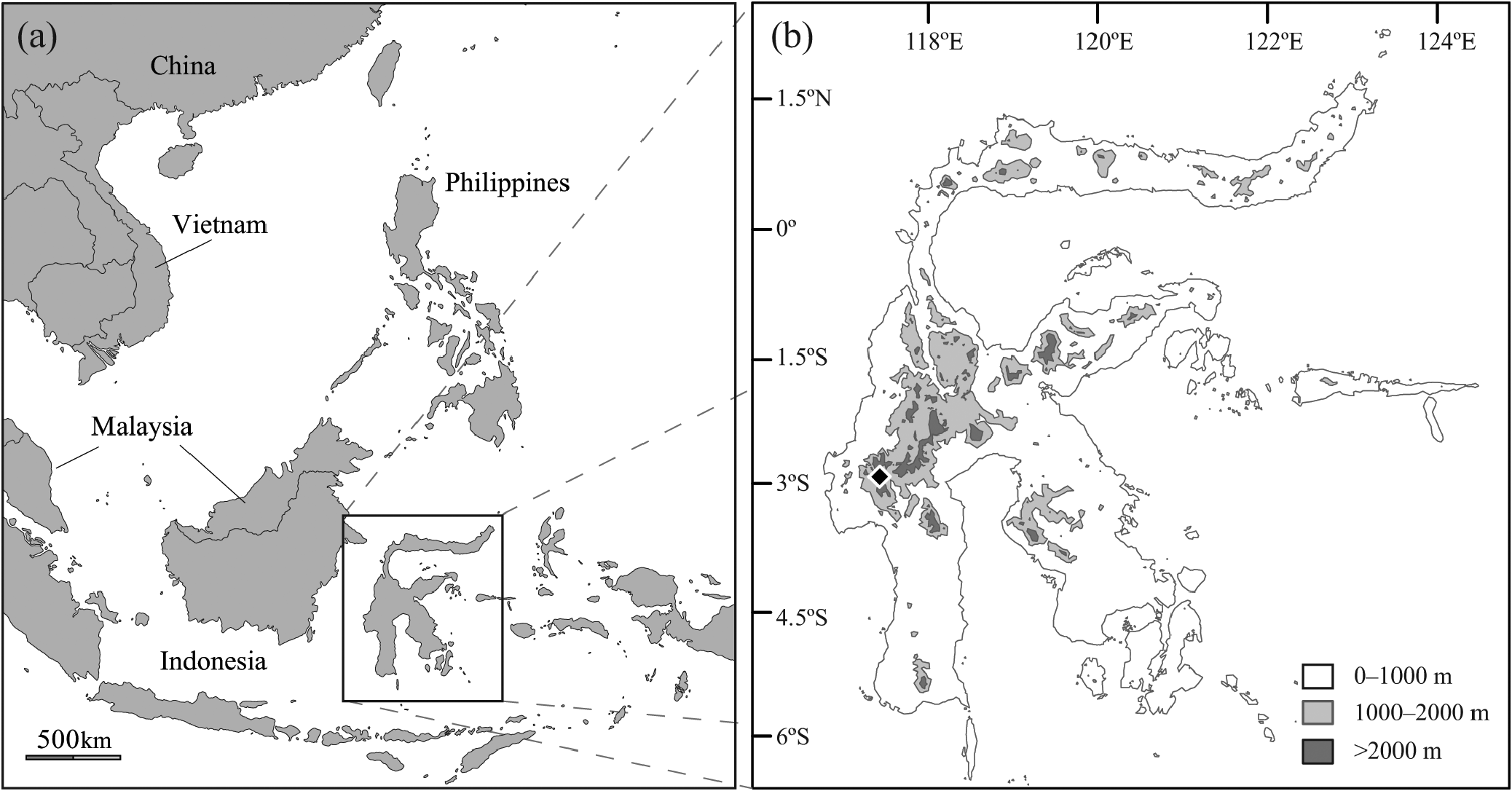
Type locality. Mount Gandangdewata (2.882898 o S, 119.386448 o E, 1571 m), Rantepangko, Mamasa, Sulawesi Barat, Indonesia ( Figure 1 View FIGURE 1 ) GoogleMaps .
Referred material. Only the holotype.
Distribution. Waiomys is known only from the type locality in lower montane rainforest of the Quarles Range of the western Sulawesi highlands.
Diagnosis. mamasae is the only known species in the genus Waiomys . Thus, generic and specific diagnoses are the same.
Etymology. The specific epiphet refers to the type locality, which is near the town of Mamasa. The local people who collected the type specimen and who had existing knowledge of the species, self-identify as Mamasan. Thus, the epithet also recognizes their knowledge and contribution to the scientific discovery of the species.
Description. Waiomys mamasae is a small, densely furred rat ( Figure 4 View FIGURE 4 ). The dorsal pelage is grey. The hairs of the underfur are <10 mm in length and have a grey base and light brown tip (~ 1–2 mm). The sparse guard hairs are grey on the basal half, dark brown on the distal half, and approximately 10 mm in length, emerging only a few millimeters above the dorsal underfur. The dorsal fur protrudes nearly vertically from the skin and can be brushed naturally in any direction. The ventral fur is as soft and dense as the dorsal underfur and of similar length with fine white guard hairs emerging <5 mm beyond the underfur. The base of the ventral underfur is dark grey whereas the distal quarter is a much paler grey. Both the dorsal and ventral coloration are relatively uniform across the head and body. The margin of the ventral fur is discrete and lies high on the body, extending from above the upper lips, at the midline of the nose, along the side of the body at the base of the limbs. A patch of ventral fur extends 15 mm up each side of the base of the tail and above the hindlimbs forming two conspicuous white spots on the rump. The dorsal fur extends to the fingers of the forefeet and to the ankles of the hindfeet.
The eyes are small (~ 2 mm diameter) and almost indistinguishable amid the fur of the head. The pinnae are grey in color, short (11 mm), heavily furred on the inner and outer surface, and nearly indistinguishable from the dorsal fur. The mystacial vibrissae are dense and stiff. A minority of the vibrissae approach 40 mm in length and extend beyond the pinnae, while the remaining whiskers are shorter at ~ 5–25 mm. The base of each vibrissa is black, but the distal quarter lacks pigmentation.
The tail is nearly 25% longer than the head and body. The ventral surface of the tail is covered in stiff white vibrissae approximately 5 mm long ( Figure 4c View FIGURE 4 ). The dorsal surface of the tail is grey-brown and similar to the color of the dorsal pelage. The tail tapers gently from the base to the tip. It is not substantially thickened or flattened and other than the ventral vibrissae, it is typical of murine rodents. The hindfeet are long (27.5% of head and body length) with short, but otherwise typical murine claws. They are light brown on the dorsal surface and dark brown on the plantar surface. The hindfeet lack the hypothenar pad ( Figure 4c View FIGURE 4 ). On the plantar surface, the first interdigital pad and thenar pad abut each other and are positioned on the lateral margin of the foot. The forefeet are small with short typical murine claws and a nailed pollex fused to the side of the palm. The palmar surface consists of three interdigital pads, a large thenar pad, and a slightly smaller hypothenar.
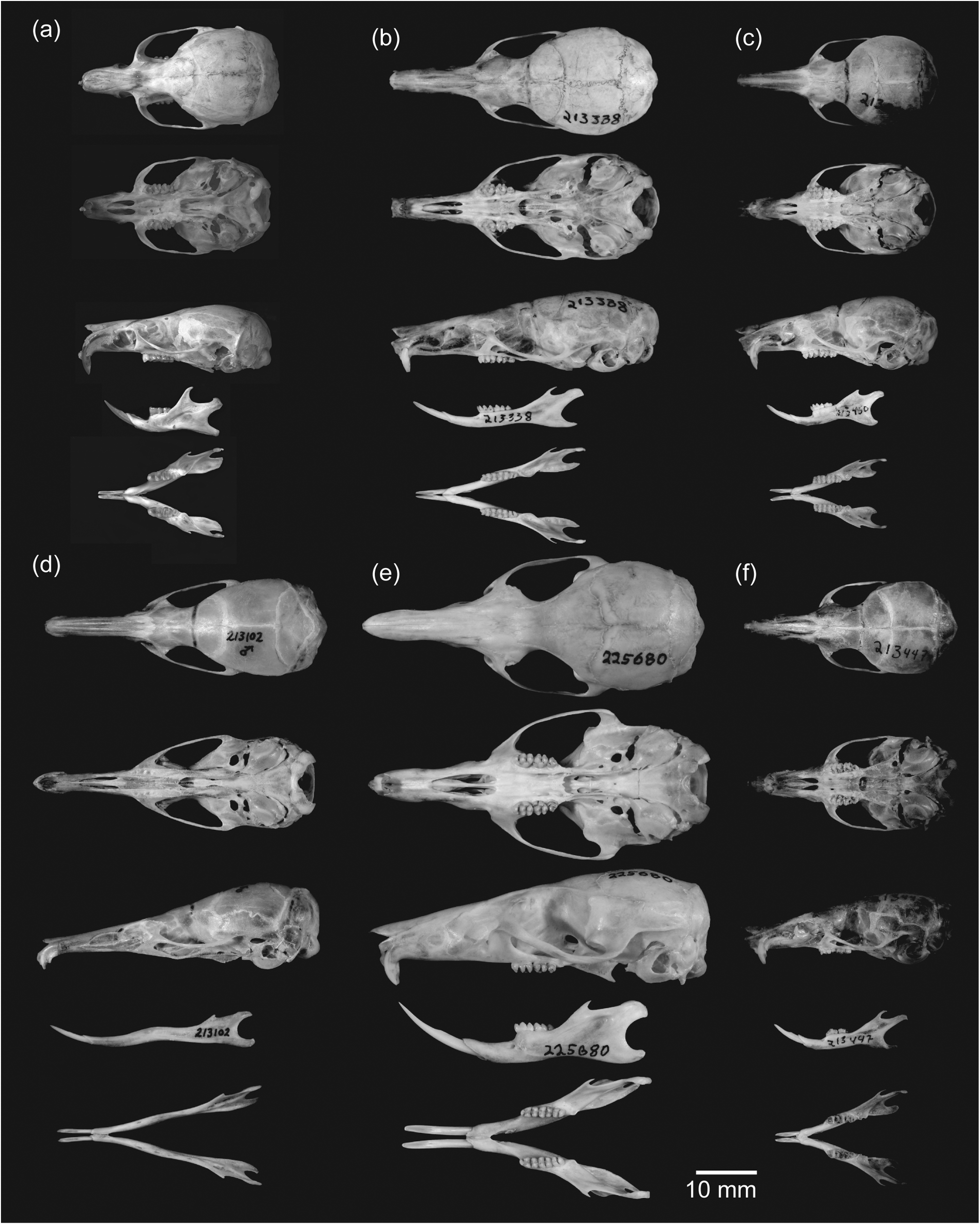
The skull of Waiomys mamasae is delicate with few prominent ridges ( Figure 5 View FIGURE 5 ). However, a distinct occipital ridge and an almost indistinguishable temporal ridge are present. For a murine, the rostrum is moderate in length relative to the basicranial region, comprising approximately one-quarter of the total length of the skull ( Table 2). The zygomatic arches are delicate and the large ovate infraorbital foramina make the zygomatic arches appear flared and squared off when viewing the skull from the dorsal or ventral aspect. The incisive foramina also are fairly broad (2.3 mm at their combined maximum breadth). The foramen magnum is large (6.17 mm in width) and equal to 42.6% of the breadth of the braincase. The occipital region is noticeably bulbous and contributes to the globose appearance of the braincase. The auditory bullae are small for a murine of this size (LB = 13% of GLS). The dentary is delicate. The angular process is narrow and elongate but otherwise the dentary is typical of murine rodents.
The upper incisors of Waiomys mamasae are narrow (each 0.5 mm wide at the root) and orthodont. The lower incisors are also narrow (0.7 mm at the base). The labial surfaces of the upper and lower incisors are pale orange. Three molars are present in both the maxillary (M1, M2 and M3) and mandibular rows (m1, m2 and m3). The occlusal surfaces of lower and upper molars form little more than basins outlining the basic murine molar cusp pattern ( Figure 5 View FIGURE 5 ) as a result of either tooth wear or an evolutionary trend toward simplification. The first and second upper molars are moderate in size for murine rodents (first upper molar 1.8 mm wide). As is the case for many murine species, the third molars, both upper and lower, are much smaller than the first and second molars.
The contents of the mouth and the nearly distended stomach of the type specimen were examined by R. Marchant (Terrestrial Invertebrates, Museum Victoria) and identified as consisting entirely of stream invertebrate larvae belonging to the families Simulidae (Black flies; genus Simulium ) and Hydropsychidae (net-spinning caddisflies; genus Cheumatopsyche ). The larvae of both genera are known to cling to the surface of stones in shallow areas of fast flowing water (Merritt & Cummins 1996), suggesting that Waiomys dives and forages for food on the stream bottom. No plant material or terrestrial animal material was identified from the mouth or stomach.
Comparisons. Waiomys mamasae is sympatric with several other murine rodents where it was collected on Mount Gandangdewata ( Figure 1 View FIGURE 1 ). These include all but two species of an endemic group of invertebrate-eating rodents on Sulawesi that are referred to as shrew rats: Melasmothrix naso , Paucidentomys vermidax , Sommeromys macrorhinos , Tateomys macrocercus , Tateomys rhinogradoides , were all detected during our surveys on Mount Gandangdewata. However, there are no records of either Echiothrix centrosa or Echiothrix leucura in the Mount Gandangdewata region. The latter species is restricted to the northern peninsula east of Gorontalo, while the former has been documented from several sites in Central and West Sulawesi (Musser & Carleton, 2005). Crunomys celebensis , another Sulawesi murine occasionally referred to as a ‘shrew rat,’ was recently shown to be closely related to species of Maxomys (Achmadi et al. 2013) and we therefore exclude it from comparisons.
In body size, Waiomys mamasae is much smaller than either species of Echiothrix , smaller than Tateomys rhinogradoides and Paucidentomys vermidax , and larger than Melasmothrix , Sommeromys , and Tateomys macrocercus . The dorsal fur of Waiomys has a similarly soft texture as that of Tateomys , but that of Waiomys is denser and paler in color. The ventral fur of Waiomys is also considerably denser and paler in color than any other Sulawesi murines. The two white patches on the rump of Waiomys are unique among murines and may represent eyespots.
Waiomys has a considerably broader and blunter rostrum than any of the shrew rats of Sulawesi. The mystacial vibrissae of Waiomys are denser and stiffer but not longer than in the shrew rats of Sulawesi. The ears of Waiomys are substantially shorter (in relative and absolute terms) and more densely furred than in any other murine on Sulawesi. The eyes of Waiomys are similar in size to those of much smaller animals, including Tateomys macrocercus and Sommeromys macrorhinos . The tail of Waiomys is similar in length and thickness to that of T. rhinogradoides , but is more sparsely haired on the dorsal surface. Among all Sulawesi murines, only Waiomys possesses ventral vibrissae on the tail. The hindfeet of Waiomys are comparable in length to the larger Tateomys rhinogradoides , but the forefeet are considerably smaller and similar in size to the feet of the smaller T. macrocercus and S. macrorhinos . The claws on the forefeet are also much smaller than in T. rhinogradoides , T. macrocercus , or Melasmothrix naso , and are comparable in size to the claws of S. macrorhinos . The position of the thenar pad and first interdigital pad on the lateral margin of the plantar surface of the hindfeet in Waiomys is distinct from those of all other murines of Sulawesi. Among Sulawesi murines, only Waiomys lacks the hypothenar pad.
(Shown are the mean, ± one standard deviation, and the range in parentheses).
Relative to its body size, the skull of Waiomys is shorter than those found in any of the shrew rats of Sulawesi and is comparable to or shorter in absolute length than the skull of the much smaller Sommeromys ( Table 2). The shortness of the skull is produced primarily by the lack of rostral elongation so prominent in the other species (rostrum is 25% of skull length compared to 35–41% in the shrew rats). However, the braincase of Waiomys also is shorter than in most of the shrew rats ( Table 2). The length of the post-palatal region in Waiomys is comparable to those of the smaller Tateomys macrocercus and only slightly larger than in Sommeromys . The zygomatic arches and braincase of Waiomys are much more square in appearance when viewed from the dorsal surface than in the other species. The infraorbital foramina are more ovate and broad in Waiomys , contributing to the flared and squared-off appearance of the zygomatic arches. The length of the auditory bulla in Waiomys is both absolutely (LB = 4.05 mm) and proportionately (LB/BBC = 0.28) shorter than in any of the shrew rats on Sulawesi (LB = 4.28-5.88 mm; LB/BBC = 0.31-0.38). Because of the trend toward elongation of the skull in the shrew rats, we used a measure of skull width (BBC) rather than of skull length (e.g. GLS) for proportional comparison. The dental formula of Waiomys is shared with all shrew rats on Sulawesi except Paucidentomys vermidax , in which the molars are absent. The upper incisors of Waiomys are both absolutely (BUI = 0.50 mm) and proportionately (BUI/BBC = 0.035) narrower than in any of the shrew rats (BUI = 0.77-1.2 mm; BUI/BBC = 0.054 -0.074). Both upper and lower incisors of Waiomys have orange enamel on the labial surface that is typical of most murines, but is absent in the shrew rats of Sulawesi. The upper molars of Waiomys are comparable in width to other shrew rats of Sulawesi (excluding Paucidentomys which lacks molars), but the lower molars are broader.
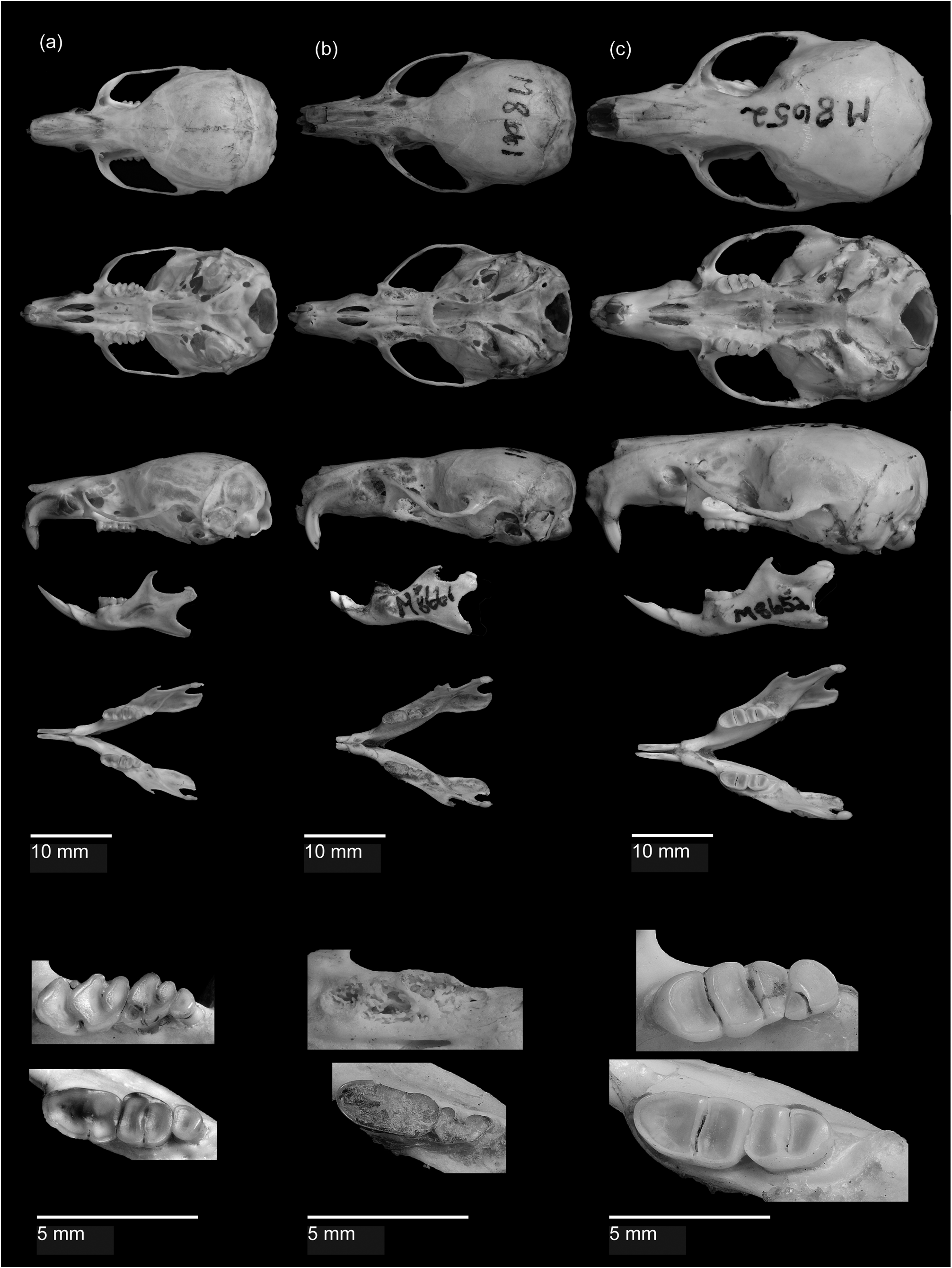
Among the water rats of New Guinea, Waiomys is similar in size to the smaller species of Hydromys ( H. hussoni and H. ziegleri ), and to the species of Baiyankamys ( B. habbema and B. shawmayeri ), but considerably smaller than Crossomys moncktoni , Hydromys chrysogaster , and Parahydromys asper ( Helgen 2005) . Like the montane species of water rats from New Guinea ( Baiyankamys and Crossomys ), Waiomys is distinguishable from the lowland water rats of New Guinea by its soft grey fur and small pinnae. Overall the fur of Waiomys is similar to that of Crossomys . However, the dorsal guard hairs of Waiomys are shorter and less prominent than those of Crossomys . The tips of the dorsal underfur in Waiomys are a paler brown than in Crossomys . The ventral underfur of Waiomys is dark grey at the base and light grey at the tip, whereas in Crossomys it is a more uniform silvery white. The dorso-ventral margin of the fur in Waiomys is similar to that of Crossomys both in its distinctiveness and its position high up on the body from the base of the nose and along the body near the base of the limbs. The white rump spots present in Waiomys are absent in Crossomys and all other New Guinea rodents. The eyes of both Waiomys and Crossomys are greatly reduced. The pinnae of Waiomys are similar in size to those of Baiyankamys but not as reduced as in Crossomys , where the pinnae are all but absent. The tail in Waiomys , Baiyankamys , and Crossomys is much longer than the head and body. Waiomys and Crossomys share the presence of stiff, white ventral vibrissae on the tail. In Waiomys , the ventral vibrissae form a single line that terminates at the base of the tail whereas in Crossomys the vibrissae bifurcate and diverge into two lines at the base of the tail. The tail of Waiomys is similar in thickness to those of terrestrial murines, but in Crossomys the tail is substantially thicker. The lack of any webbing or hairs on the margins of the plantar surface of Waiomys distinguishes it from all water rats of New Guinea. In both Waiomys and Crossomys , the hypothenar pad is absent from the plantar surface of the hindfeet and the thenar pad is positioned on the lateral margin of the foot. Like Crossomys and Baiyankamys , the forelimbs of Waiomys are small.
The skull of Waiomys is similar in size and robustness to those of Baiyankamys and Hydromys hussoni ( Table 3, Figure 6 View FIGURE 6 ; H. hussoni comparison in Helgen 2005). Waiomys shares some cranial features with Baiyankamys and Crossomys , the montane water rats of New Guinea ( Helgen 2005), to the exclusion of the lowland water rats of New Guinea. These include (1) slender and delicate zygomata that curve downward from the maxillary roots to the squamosal roots; (2) a tapering rostrum; (3) extremely narrow incisors; (4) a mesopterygoid fossa that is narrower than the width between the lingual margins of the first upper molars. In addition, two morphological characters clearly distinguish Waiomys from all water rats of New Guinea: (1) the presence in Waiomys of the third mandibular molar and (2) the absence in Waiomys of the masseteric process of the zygomatic plate.
(Shown are the mean, ± one standard deviation, and the range in parentheses).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.