
Micromaldane shackletoni, Darbyshire, Teresa, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3683.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:0CDAAE0B-E1B2-4D42-8C09-E07534EF47EE |
|
DOI |
https://doi.org/10.5281/zenodo.5664906 |
|
persistent identifier |
https://treatment.plazi.org/id/F66815EB-EBC6-48F7-A186-2A9093C4F32C |
|
taxon LSID |
lsid:zoobank.org:act:F66815EB-EBC6-48F7-A186-2A9093C4F32C |
|
treatment provided by |
Plazi |
|
scientific name |
Micromaldane shackletoni |
| status |
sp. nov. |
Micromaldane shackletoni View in CoL n. sp.
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Table 1 View TABLE 1
Material examined. SW Atlantic, Falkland Islands: Cochon Island, Stn 14 (51° 36.217ʹ S, 057° 47.585ʹ W), rock scraping, 10.4 m, holotype ( NMW.Z.2011.039.0001), 27 paratypes (1—AM W.43557; 2— NHMUK 2013.416; 24— NMW.Z.2011.039.0002–0024; NMW.Z.2011.039.0025 on SEM stub), 25.11.2011; Cochon Island, Stn 10 (51° 36.287ʹ S, 057° 47.684ʹ W), rock scraping, 9.5 m, 1 paratype ( NMW.Z.2011.039.0026), 24.11.2011; Cochon Island, Stn 11 (51° 36.377ʹ S, 057° 47.489ʹ W), from Chaetopterus tube, 9.6 m, 1 paratype ( NMW.Z.2011.039.0027), 24.11.2011; Cochon Island, Stn 13 (51° 36.322ʹ S, 057° 47.132ʹ W), bryozoan scraping, 13.6 m, 3 paratypes ( NMW.Z.2011.039.0028–0030), 25.11.2011; Cochon Island, Stn 16B (51° 36.366ʹS, 057° 47.082ʹW), hydroid/bryozoan turf scraping, 12.5 m, 62 paratypes (1—AM W.43558; 51— NMW.Z.2011.039.0031–0081; NMW.Z.2011.039.0082–0087 on SEM stubs; 2— USNM 1207943; 2— ZMH P- 27462), 26.11.2011.
Description. Holotype ( Fig. 1 View FIGURE 1 A) complete, 8.3 mm long, 0.3 mm wide for 21 chaetigers. Complete paratypes 3.6–11.0 mm long, 0.3 mm wide for 19–23 chaetigers. The following description is based on preserved specimens with 19 or more chaetigers.
Body cylindrical, slightly wider on prostomial segment but uniform in width for rest of body. Body cream or yellowish in alcohol, some brown pigment visible on prostomial and anterior two segments; not observed alive. Prostomium fused to peristomium, rounded anteriorly, forming a slight arched dorsal keel. Small cluster of black eyespots visible each side of prostomium anterior to buccal opening. Nuchal organs rounded ciliated pits with small, central groove anterior to each ( Fig. 1 View FIGURE 1 B).
Notochaetae of two types: straight (under light microscopy, tips curved under SEM conditions), stout, lancetshaped chaetae ( Fig. 1 View FIGURE 1 C, D), distally tapering with covering of fine hairs, and fine capillary chaetae with similar haired covering ( Fig. 1 View FIGURE 1 D). All chaetigers with 1–2 (rarely 3) lancet-shaped chaetae and 1–3 capillary chaetae.
Neuropodia of all chaetigers with uncini, up to 9 (rarely 10) in a single row. Chaetiger 1 with 1 or 2 uncini increasing to maximum number (4–10, number highly variable) on chaetigers 9–11, thereafter decreasing to 1 or 2 on final chaetiger ( Fig. 2 View FIGURE 2 ). Uncini strongly curved with long shafts, large main fang with 4 smaller teeth above and numerous additional teeth surrounding ( Fig. 1 View FIGURE 1 E). To each side of main fang, subrostral barbules project upwards and forwards.
Chaetigers increase in length to median chaetigers then decrease progressively thereafter. Single pre-anal achaetigerous segment present. Anal plaque funnel-shaped, radially symmetrical, edge gently crenated (number variable); anus central ( Fig. 1 View FIGURE 1 F).
Tube soft, composed of mucus and loosely bound sand grains. Numerous individuals identified with both eggs and sperm ( Fig. 3 View FIGURE 3 A) floating free in the coelomic cavity and some tubes found with directly developing larvae (see below: Reproduction). Larvae were at different stages from pre-chaetigerous to 14 chaetigers ( Fig. 3 View FIGURE 3 B–G). Spermathecae absent.
Methyl green staining produced ‘speckles’ over the entire body, more ventral than dorsal, with staining more concentrated on ventral side of anterior 6 chaetigers, prostomium (ventral and anterior dorsal) and pre-anal chaetiger (dorsal and ventral).
Variation. The number of chaetigers is very variable with independent (having their own tube) individuals ranging in size from 15 to 23 chaetigers. Most specimens with 19 chaetigers or more had internal reproductive products visible while only a small number of 18-chaetiger specimens, none smaller, were found containing eggs and sperm. The latter is therefore taken as the size of onset of maturity. Only a single individual was found with 23 chaetigers, with the majority of mature specimens identified having 19 or 20.
Length was extremely variable even among those specimens with the same number of chaetigers (none of those individuals measured showed obvious signs of regeneration). Body width showed little variation regardless of length.
The number of uncini on each chaetiger increased with the total number of segments but showed large variation even within the same size category. Fig. 2 View FIGURE 2 compares the average number of uncini per segment for each of the mature adult size groups (18–23 chaetigers). Average chaetal counts are highest in the mid-body region (chaetigers 8–11) where a general increasing trend relative to total chaetiger number is evident.
Reproduction. Eggs and sperm were both visible within reproductive individuals and were confirmed as such using scanning electron microscopy ( Fig. 3 View FIGURE 3 A: egg 30 µm diameter, sperm ‘packet’ 30x16 µm). As no specimens contained only sperm or only eggs, and no spermathecae were found, the species is presumed to be a simultaneous hermaphrodite as described previously for M. androgyne Rouse, 1990 . More advanced individuals contained larger pre-chaetigerous embryos and a single animal was found with a large swelling on one side of the body that appeared to be the ‘budding-off’ of ripe embryos ( Fig. 3 View FIGURE 3 B). Additional animals were found with the prechaetigerous larvae (up to 12) inside the tube rather than the body ( Fig. 3 View FIGURE 3 C). Further specimens were identified with 5–7 chaetiger or 13–14 chaetiger larvae still developing inside the parental tube ( Fig. 3 View FIGURE 3 D). In each case, all larvae within a single tube were at a similar stage of development. The number of developing larvae reduced with increasing size from up to 12 pre-chaetigerous larvae in a tube to only three 13 or 14–chaetiger larvae. These observations, however, are only based on a very small number of specimens.
Larval development was investigated in those larvae found in parental tubes. No cilia were visible on any of the larvae at any stage ( Fig. 3 View FIGURE 3 E–G). Early stage larvae, with no differentiation visible on the body, were approximately 210 µm by 160 µm in size ( Fig. 3 View FIGURE 3 E). By the 7–chaetiger stage ( Fig. 3 View FIGURE 3 F: 0.58 mm long, 0.1 mm wide) segmentation was visible with an identifiable prostomial segment and associated buccal opening. No eyespots were visible at this stage. Notochaetae of both types were present but uncini were absent and the posterior end was still undeveloped. At 14 chaetigers ( Fig. 3 View FIGURE 3 G: 0.84 mm long, 0.1 mm wide), just prior to leaving the parental tube, eyespots were visible on the prostomium. Fig. 3 View FIGURE 3 D shows a 14–chaetiger larva positioned alongside but in the opposing direction to the parental body as indicated by the visible eyespots of both animals (see arrows on figure). Notochaetae had developed on all visible segments and uncini on all but the most posterior chaetiger. The anus was visible but the anal plaque was still undeveloped. Investigation of larger individuals showed that this character does not fully develop until around the 16–chaetiger stage. At 15 chaetigers in size, animals were independent and had constructed their own tube.
Etymology. This species is named after Sir Ernest Shackleton, Antarctic explorer, for whom the Shackleton Scholarship Fund is commemorated, in recognition of the Fund’s support of this work.
Habitat. All specimens were collected subtidally (9–14 m) from rock scrapings of epifaunal ‘turf’. Tubes were attached to any available structure within the ‘turf’ including hydroids, bryozoans and polychaete tubes (including those of other Micromaldane ).
Remarks. Micromaldane shackletoni n. sp. is distinguishable from most other species of Micromaldane by having both a pre-anal achaetigerous segment and straight lancet chaetae ( Table 1 View TABLE 1 ). This combination of characters is closest to M. androgyne and M. bispinosa . However, M. androgyne has only single or no capillary chaetae in each chaetiger, as opposed to 1–3 in M. shackletoni n. sp. and straight nuchal grooves as opposed to rounded, ciliated pits. For a smaller number of segments (maximum 23 as opposed to maximum 25), M. shackletoni n. sp. is also a much longer animal, almost twice the length, than M. androgyne . A comparison of larval development between the two species further confirms the distinction between them: larvae of M. androgyne remain in the parental tube until 19 chaetigers as opposed to 14 chaetigers for larvae of M. shackletoni n. sp.; uncini are present on most chaetigers by the 14–chaetiger stage in M. shackletoni n. sp. but not until 19 chaetigers in M. androgyne and development of the anal plaque is earlier in M. androgyne (visible from 11 chaetigers) than in M. shackletoni n. sp. where the anal plaque is still undeveloped at 14 chaetigers. Micromaldane bispinosa remains unique in the group in possessing two types of uncini whereas M. shackletoni n. sp., like the rest, has only one type.
Past suggestions that the genus Micromaldane represents only a post-larval stage of another maldanid have been shown to be incorrect by the taxonomic and reproductive studies carried out by Rouse (1990; 1992a – b), in which several different species of Micromaldane were shown to be fully mature, reproductive adults. Both gonochorism ( M. nutricula , M. pamelae , M. rubrospermatheca ) and hermaphroditism ( M. androgyne ) are represented in the genus ( Rouse 1990, 1992a), and all brood directly with larvae developing within the adult tube. No reproductive details were given for M. jonesi ( Kumaraswamy Achari 1968) and there were only brief mentions of the presence of eggs in M. ornithochaeta ( Mesnil 1897) and possible sperm in M. bispinosa by Hartmann- Schröder (1960).
Larval development of Micromaldane species was compared with that of other maldanid genera by Rouse (1992a). Within the Maldanidae , only Euclymene oerstedii ( Claparède, 1863) has been reported to show hermaphroditism, although only a minority of the population exhibited this, the rest being gonochoristic ( Pilgrim 1964). Boguea enigmatica Hartman, 1945 is the only other maldanid known to exhibit direct development similar to that of Micromaldane ( Rouse 1992a) . However, larvae of that species were found at several stages of development within a single tube and hermaphroditism was not evident ( Wolf 1983).
Micromaldane shackletoni n. sp. represents only the second report of simultaneous hermaphroditism in the genus, third in the family as a whole, thereby providing further evidence for the validity of the genus.
This work was funded by both the Shackleton Scholarship Fund ( Falkland Islands) and Amgueddfa Cymru – National Museum Wales. Additional logistical support was provided by the Fisheries Department of the Falkland Islands, the Shallow Marine Surveys Group, with whom all diving was undertaken, and the South Atlantic Environmental Research Institute (SAERI).
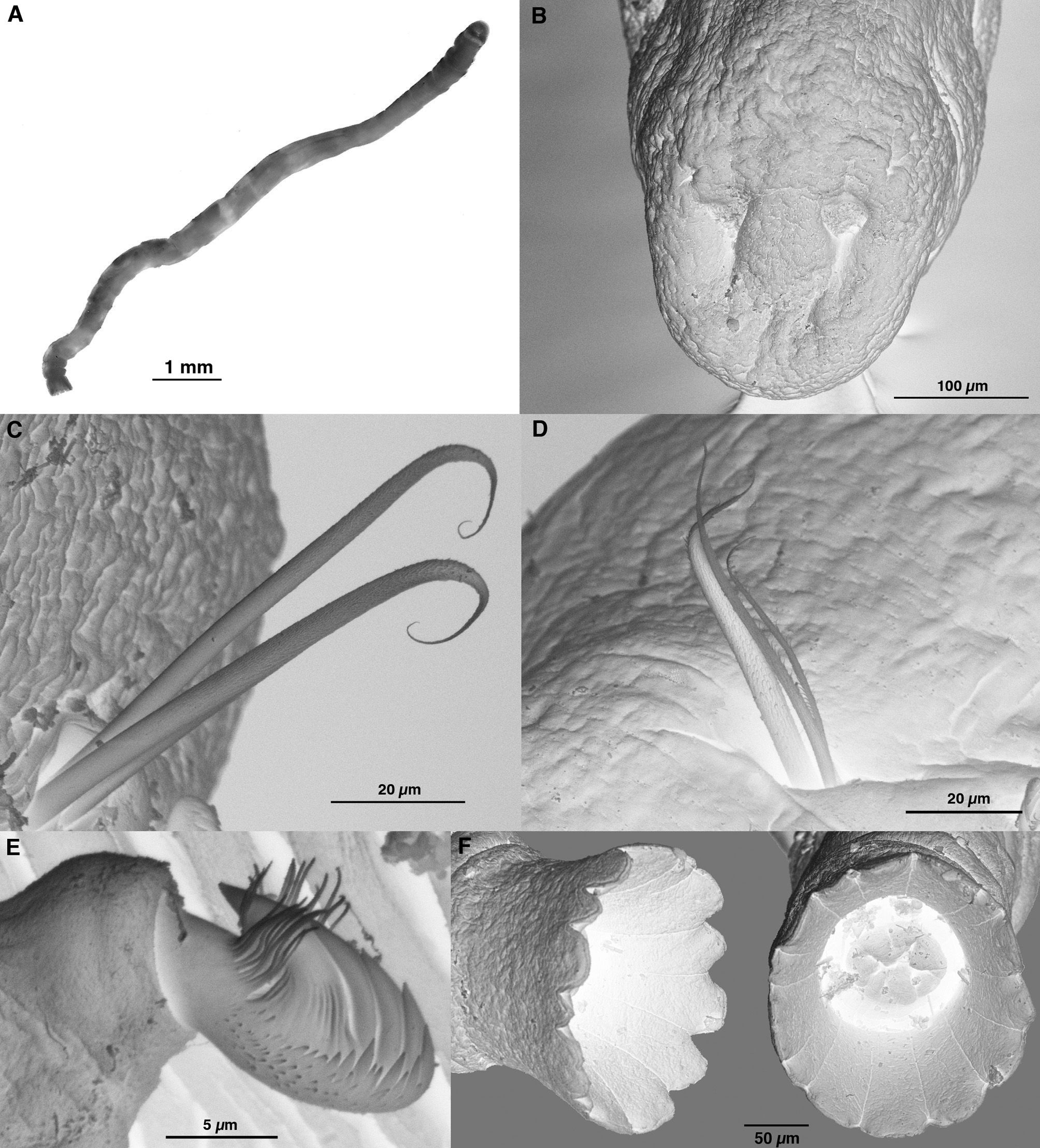
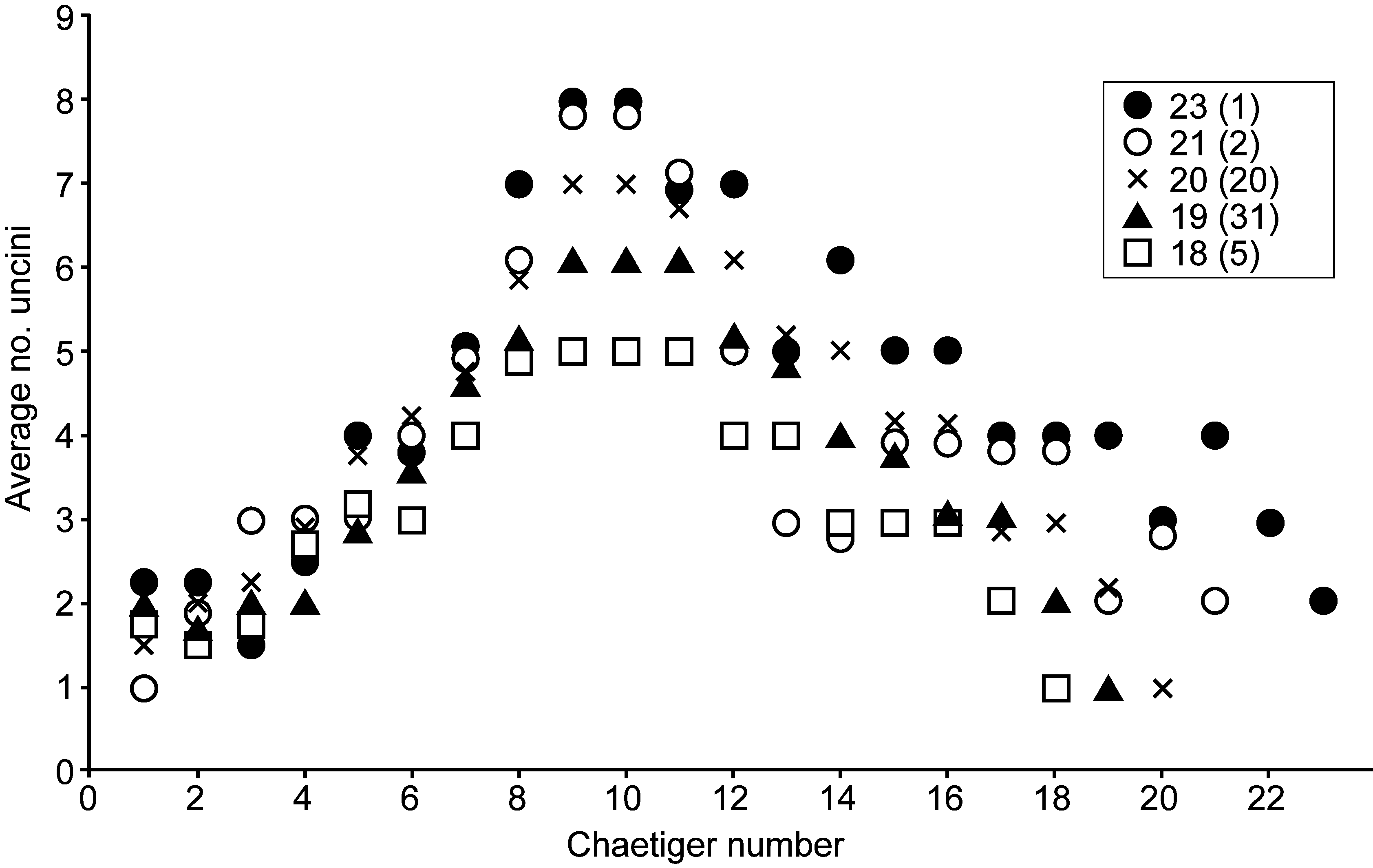
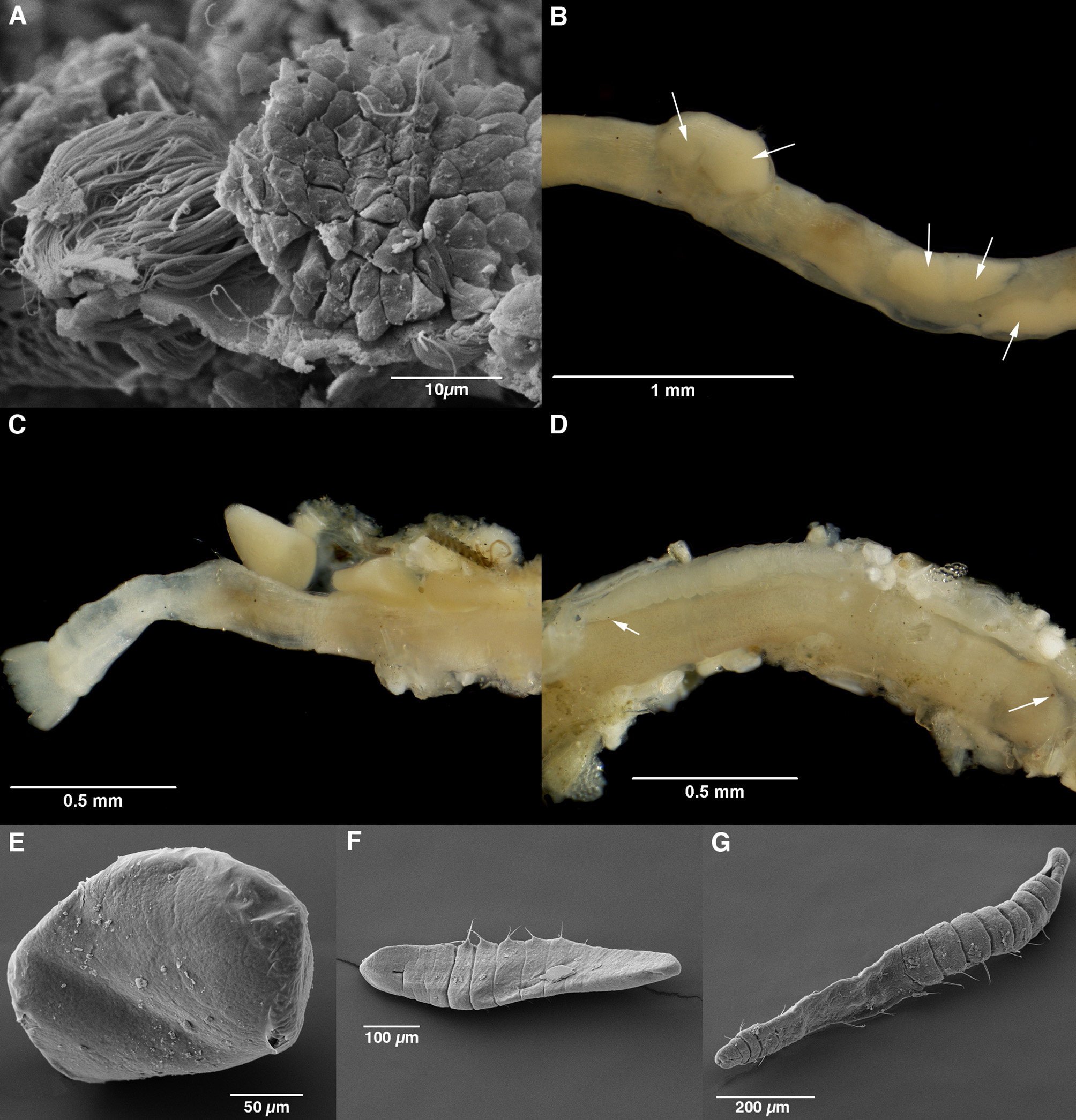
TABLE 1. Comparison of morphological characters for all currently described species of Micromaldane.
| Species M. androgyne Rouse, 1990 | Type Locality Australia | Max segments 25 | Achaetous pre- anal segment? Y | No. capillaries 0–1 | No. & form of lancet chaetae 1–2 straight |
|---|---|---|---|---|---|
| M. bispinosa Hartmann-Schröder, 1960 | Red Sea | 21 | Y | 1–3 | 1–2 straight |
| M. jonesi Kumaraswamy Achari, 1968 | India | 19 | N | absent | ? spatulate |
| M. nutricula Rouse, 1990 | Australia | 19 | N | 0–? | 1–3 straight |
| M. ornithochaeta Mesnil, 1897 | France | 17 | Y | 2 | 1–2 spatulate |
| M. pamelae Rouse, 1990 | Australia | 23 | N | 1–3 | 1–3 straight |
| M. rubrospermatheca Rouse, 1990 | Australia | 21 | N | 0–2 | 1 straight |
| M. shackletoni n. sp. Darbyshire, 2013 | Falkland Islands | 23 | Y | 1–3 | 1–2 straight |
| TABLE 1. (Ccontinued) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.