
Eurysilenium truncatum M. Sars, 1870
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4579.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A4015309-D9B3-4BB7-ABCB-B88A1F8CE5FC |
|
persistent identifier |
https://treatment.plazi.org/id/97720E2D-FFFF-D60A-CBF7-BAB50163F05A |
|
treatment provided by |
Plazi |
|
scientific name |
Eurysilenium truncatum M. Sars, 1870 |
| status |
|
Eurysilenium truncatum M. Sars, 1870
Scandinavian material examined: 3♀♀ from Eucranta villosa Malmgren, 1866 , Snorre TLP/UPA, Stn 24-5 (61.48564°N, 02.231242°E), depth 332 m, 0 7 June 2005; collected by A. Sikorski; NHMUK GoogleMaps Reg. No. 2013.147- 148 , 1♀ used for SEM. 1♀ from E. villosa , 3940 Arenaria 2, Stn 1-4 (72.63017°N, 24.92828°E), depth 270 m, 24 June 2007; collected by A. Sikorski; NHMUK GoogleMaps Reg. No. 2015.2998 . 1♀ from E. villosa, Snøhvit N, Stn 13-2 (71.49036°N, 21.07921°E), depth 329 m, 19 June 2007; collected by A. Sikorski; NHMUK GoogleMaps Reg. No. 2015.2999 . 1♀ from Eunone sp., Troll B, Stn 44-4 (60.7108°N, 03.5100671°E), depth 322 m, 27 May 2001; collected by A. Sikorski; NHMUK GoogleMaps Reg. No. 2015.3000 . 3♀♀ (2 ovigerous) from G. cirrhosa , SW of Yttre Vattenholmen , Kosterfjord, Sweden, depth unknown, 27 August 1986; collected by A. Mackie. Deposited National Museum of Wales, Reg No. NMWZ 1986.108 View Materials .44. 1 ovigerous ♀ from G. cirrhosa , SW of Yttre Vattenholm , Kosterfjord, Sweden, depth 60-80 m, 28 October 1989; collected by A. Mackie, Deposited National Museum of Wales, Reg No. NMWZ .1989.125.8.
British material examined: 1 ovigerous ♀ from G. cirrhosa , East Shetland Basin, Stn 90, (60 o 34.49’N, 0 1 o 21.33’W) depth 136 m, 25 July 2007; collected by P.R. Garwood. NHMUK Reg. No. 2017.479. 1 mature ♀ and 5 juvenile ♀ attached to single G. cirrhosa, Isle of Mull, Loch Scridain , SEPA Stn 22 (55 o 21.14’N, 0 6 o 59.53’W), depth GoogleMaps 26 m, 24 May 2016. NHMUK Reg. No. 2018.134 .
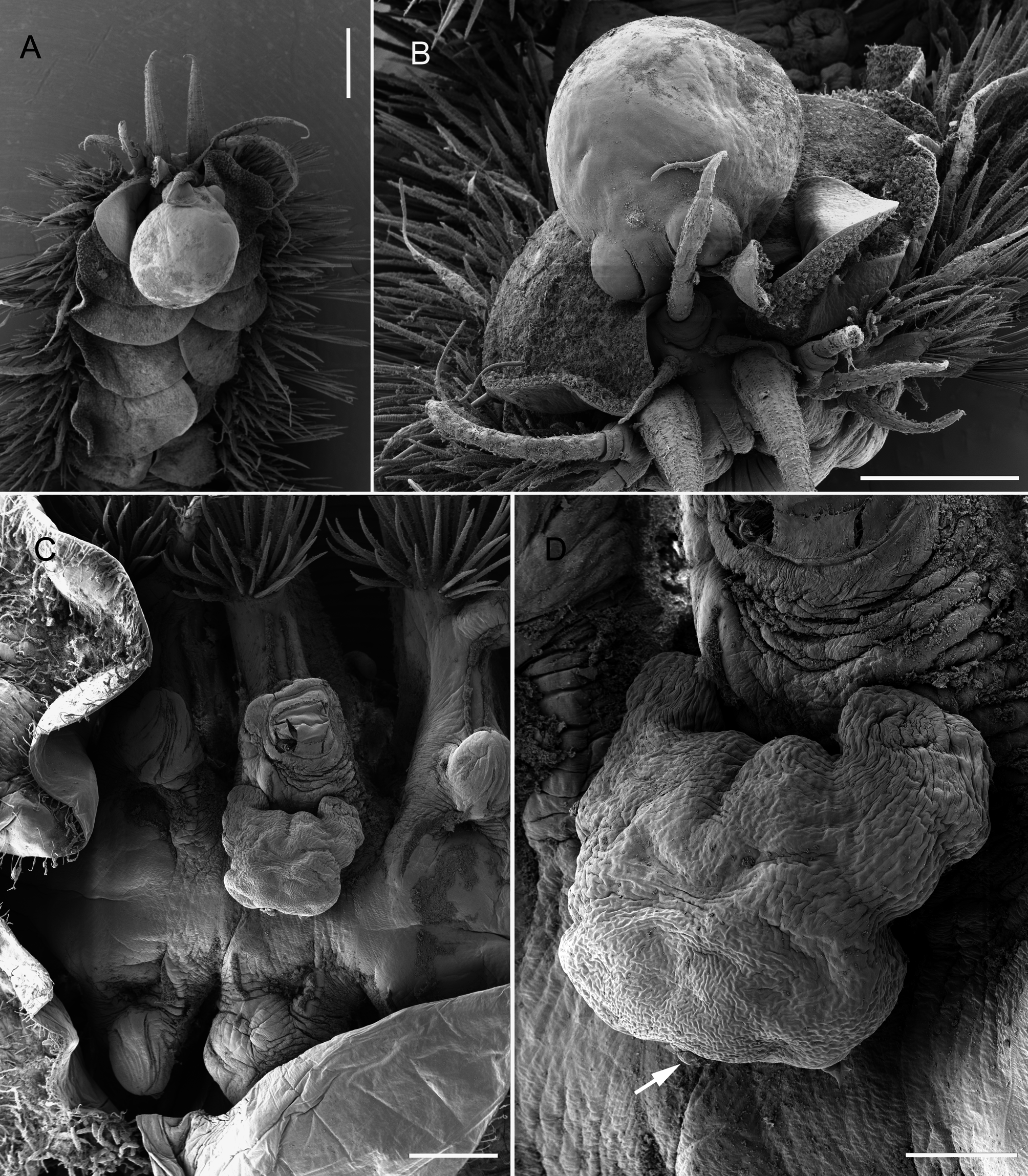
Differential diagnosis. Ectosoma of adult female subrectangular in outline, with anterior margin produced anterolaterally into paired lobate shoulders ( Figs 7C, D View FIGURE 7 , 8B, C View FIGURE 8 ). Lateral margins irregular, tapering slightly posteriorly ( Figs 7D View FIGURE 7 , 8C View FIGURE 8 ). Length of ectosoma ranging from 0.73 mm to 1.2 mm, and width ranging from 0.80 mm to 1.48 mm. Posterior margin of ectosoma straight to weakly rounded. Ectosoma dorsoventrally flattened, with paired genital apertures located ventrally close to posterior margin and either side of mid-line. Genital apertures slit-like, with sclerotized opercular plates anterior and posterior to slit ( Fig. 8C View FIGURE 8 ). Egg sacs curved (reniform), directed laterally, eggs multiseriate. Stalk originating from flat underside in midline and in anterior quarter to third of ectosoma. Endosoma ( Fig. 8A View FIGURE 8 ) elongate, typically 5 to 6 mm in length in adult female, circular in cross-section and tapering distally, terminating in globular swelling; typically looping around gut.
Male ( Fig. 8E View FIGURE 8 ) about 250 µm in total body length excluding caudal rami; body indistinctly segmented, tapering posteriorly, comprising well-defined, subrectangular cephalothorax and indistinctly 4- or 5-segmented postcephalothoracic trunk. Penultimate trunk somite with paired lobate appendages located ventrally. Anal somite bearing paired elongate caudal rami, each distinctly hooked at tip; rounded lobes present lateral to bases of caudal rami. Cephalothorax attached to female by ventral surface. Single pair of stylet-like structures present, either side of midline, in oral region anterior to mid-length of cephalothorax.
Remarks. The original description of the male of E. truncatum suggested that the male was attached to the female by a sticky secretion and no mouthparts were noted ( M. Sars 1870; Lützen 1964a). The male of E. truncatum described here was attached to the female between the genital apertures. The attachment interface was the ventral surface of the cephalothorax and a secretion may have been involved, but paired stylets were also present in the oral region, as described for E. oblongum by Lützen (1964a). These stylets presumably represent modified mouthparts. The body length of the male given by Lützen (1964a) was 250 µm excluding the caudal rami. López-González et al. (2006) described male E. australis as possessing an elongate body, tapering posteriorly, indistinctly segmented, limbless, and with hooked caudal rami posteriorly. On the ventral surface of the cephalothorax they described two “pairs of denticulated triangular elements”, a “pair of marginal bifid elements” and a “pair of tubular flaccid tubes that could be interpreted as spermiducts (?)”, plus a pecten-like area of uncertain homology in the mid-line. López-González et al. (2006) reported more paired structures than any previous study but did not offer any interpretations of the homology of these structures. As a generalization, it appears that males of Eurysilenium attach to the female by the ventral surface of the cephalothorax, and that this surface is provided with vestiges of at least one pair of appendages. López-González et al. (2006) interpreted the posterior-most pair of these structures (the flaccid tubes) as “spermiducts”. Presumably they are suggesting that these ducts represent spermatophore tubules through which the contents of the spermatophores will eventually discharge, as described for Herpyllobius males by Lützen (1966). López-González et al. (2006) similarly described paired pores that they attributed to the “spermiducts”, in the scar where the cephalothorax of the male of Gottoniella antarctica López- González, Bresciani & Conradi, 2006 was attached to the female. This implies that males of Eurysilenium and Gottoniella discharge their spermatophore contents anteriorly via their cephalothoracic attachment onto the female, as in Herpyllobius , rather than posteriorly via paired genital apertures on the urosome, as is typical of the vast majority of copepods.
The original host of E. truncatum listed by Sars (1870) was Harmothoe imbricata (as Polynoe cirrata ), but according to Lützen (1964a) the most common host is Gattyana cirrhosa . Lützen (1964a) also noted that E. truncatum was invariably attached to the dorsal surface of the host in mid-body, typically between setigers 8 and 15. The three adult females reported from a single individual of Eucranta villosa (collected off the coast of southern Norway) were attached on setigers 14, 18 ( Fig. 8C View FIGURE 8 ) and 25 of a single host. The parasites are typically concealed beneath the elytra of the host.
This species has been reported from numerous locations around southern Norway and into the Skagerrak and Kattegat ( Lützen 1964a), and from Scotland, off Shetland and in the Firth of Clyde in Scotland ( O’Reilly et al. 2011). The depth range recorded in the present account for material of this species from Scandinavian waters was 270 to 332 m whereas in Scottish waters it occurs as shallow as 26– 80 m. The record here of E. truncatum on Eucranta villosa collected at 72.6°N is the most northerly record to date and is the first from this host.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |