
Melinnacheres ergasiloides M. Sars, 1870
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4579.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A4015309-D9B3-4BB7-ABCB-B88A1F8CE5FC |
|
DOI |
https://doi.org/10.5281/zenodo.5927038 |
|
persistent identifier |
https://treatment.plazi.org/id/97720E2D-FFCA-D638-CBF7-BC6807A2F3F0 |
|
treatment provided by |
Plazi |
|
scientific name |
Melinnacheres ergasiloides M. Sars, 1870 |
| status |
|
Melinnacheres ergasiloides M. Sars, 1870 View in CoL
Material examined: 3 ovigerous ♀♀ from Melinna elisabethae McIntosh, 1914 , Resi Stagnes, Stn 5-2 (68° 48.459’N, 16° 36.753’E), depth 74 m, 28 May 2006; collected by A. Sikorski; NHMUK Reg. Nos 2015.3004-3006. 1 ovigerous ♀ from M. elisabethae, Argus Miljø, Stn Ø 2-2 (67° 14.692’’N, 15° 23.094’E), depth 145 m, 2009; collected by A. Sikorski GoogleMaps ; NHMUK Reg. No. 2015.3007. 1 ovigerous ♀ from M. elisabethae, Norvarg, Stn 1-4 (72.88696°’N, 25.79947°E), depth 371 m, 29 May 2010; collected by A. Sikorski ; NHMUK Reg. No. 2015.3008 .
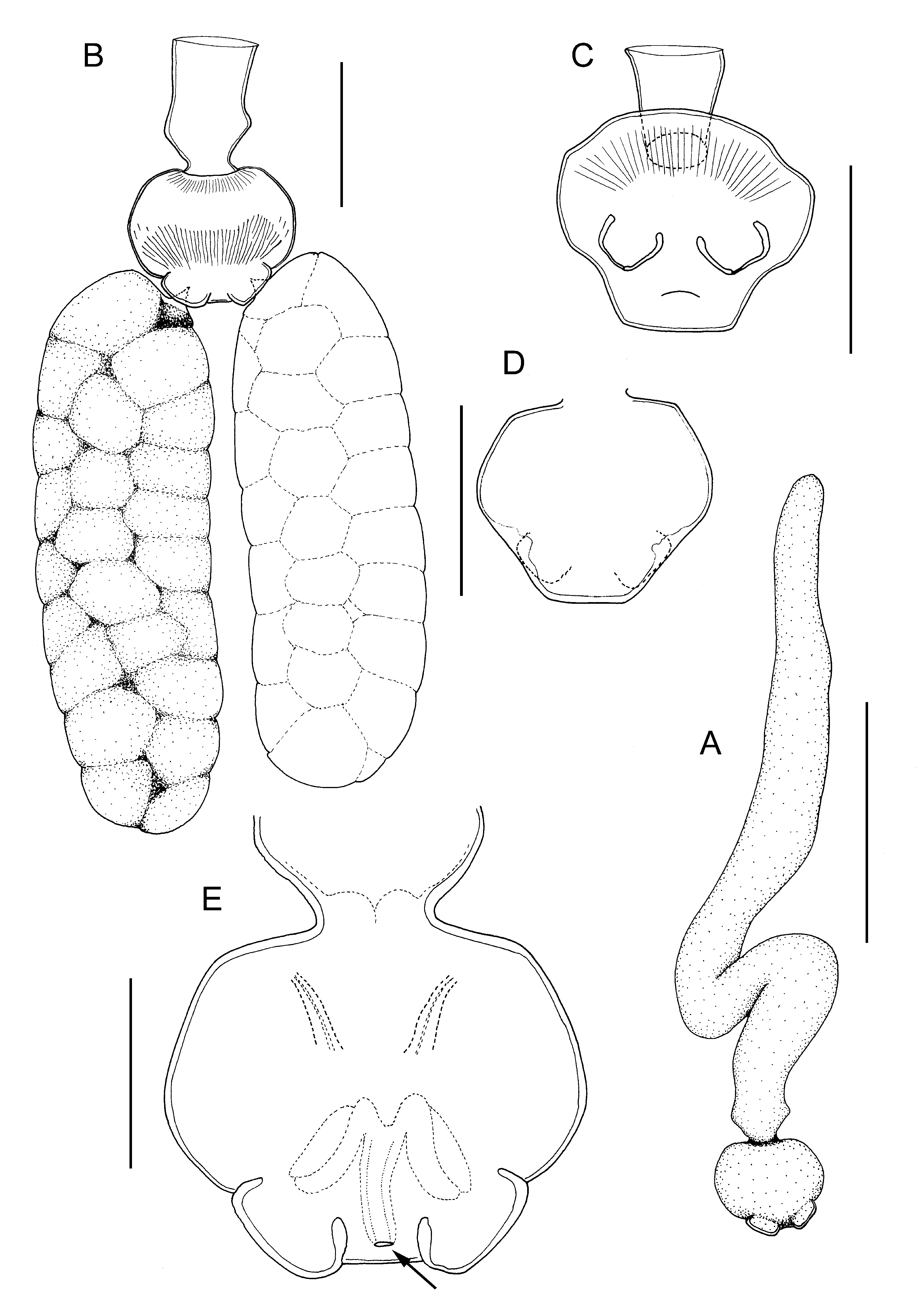
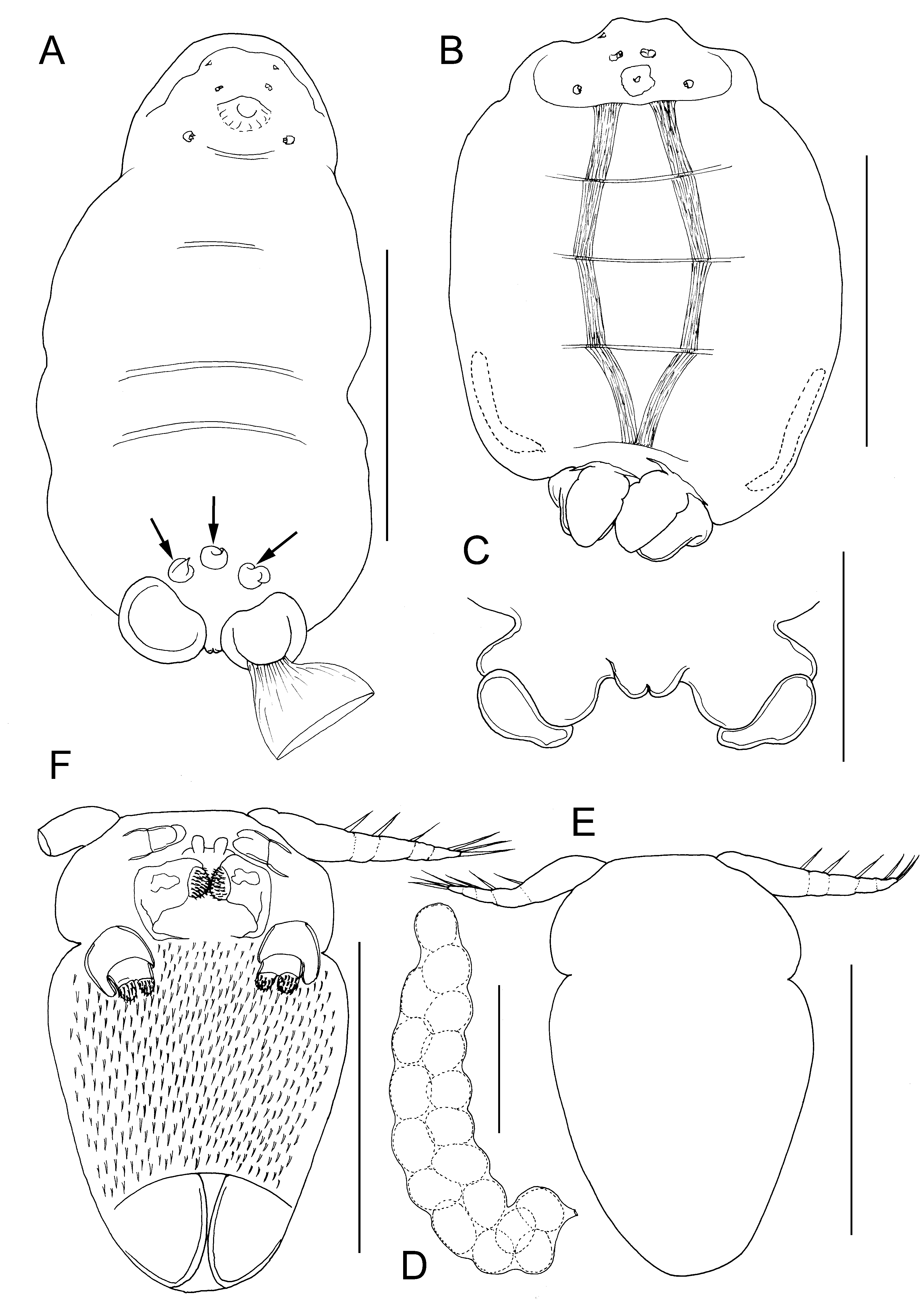
Differential diagnosis. Body of adult female ovoid in outline ( Fig. 14A, B View FIGURE 14 ), dorsoventrally flattened; mean body length 0.99 mm (range 0.93 to 1.08 mm, based on 5 ovigerous specimens); body from 1.38 to 1.94 times longer than wide (mean = 1.6 times) according to state of contraction (cf. Fig. 14A, B View FIGURE 14 ). Ventral longitudinal trunk muscles visible through ventral body wall and transverse internal sutures also visible, marking planes of former inter-segmental boundaries and providing attachment sites for longitudinal trunk muscles ( Fig. 14B View FIGURE 14 ). Paired genital apertures carried on conspicuous genital swellings located ventrally near posterior margin. Small anal prominence with median anal slit, present dorsally above mid-posterior margin ( Fig. 14C View FIGURE 14 ). Female attached to host via short stalk located anteriorly on ventral surface, leading to short bulla (endosoma) inserted in host. Paired antennules, antennae and maxillae present around base of stalk ( Fig. 14A View FIGURE 14 ). Antennules short, tapering distally, indistinctly subdivided into broader basal segment and tapering distal segment armed with apical seta. Antennae located anterior to stalk, orientated transversely with apex directed towards midline: 2-segmented, basal segment unarmed, apical segment with paired distal adhesion pads. Maxillae located posterior to stalk; comprising robust basal segment with inner distal corner produced into spinous process; terminal segment bearing 2 adhesion pads. Cement glands short, extending only 30% of length of trunk, lying within zone derived from fourth pedigerous somite ( Fig. 14B View FIGURE 14 ). Egg sacs weakly curved, irregularly biseriate ( Fig. 14D View FIGURE 14 ).
Adult male body ovoid, indistinctly divided into anterior cephalothorax and tapering posterior trunk ( Fig. 14E, F View FIGURE 14 ); body length about 155 µm, maximum width about 100 µm, at anterior part of trunk. Cephalothoracic region bearing paired antennules, antennae and maxillae surrounding oral region. Trunk lacking defined limbs. Ventral surface of trunk ornamented with dense covering of fine spinules ( Fig. 14F View FIGURE 14 ). Antennules elongate, tapering towards tip, indistinctly segmented; armed with 3 setae along anterior margin and 3 setal elements around tip ( Fig. 14E, F View FIGURE 14 ). Antennae 2-segmented ( Fig. 14F View FIGURE 14 ); basal segment unarmed, apical segment with 2 rounded apical pads. Paired labral processes present either side of ventral midline, just anterior to chitinous oral framework. Oral region defined by sub-rectangular framework of chitinous thickenings enclosing paired, medially-directed, rounded spinose pads ( Fig. 14F View FIGURE 14 ), interpreted as mandibles by Bresciani & Lützen (1975). Maxilla ( Fig. 12F View FIGURE 12 ) comprising robust basal segment with inner distal corner produced into blunt-tipped spinous process; distal segment bearing 2 adhesion pads ornamented with dense covering of fine spinules.
Remarks. Melinnacheres ergasiloides was originally reported from the ampharetid worm Melinna cristata ( M. Sars, 1870) caught in Oslofjord in southern Norway ( M. Sars 1870). Sars described and illustrated the external morphology of both sexes in detail. Over a century later, Bresciani & Lützen (1975) redescribed both sexes on the basis of new material collected from the same host, M. cristata , caught off the western Swedish coast in the Skagerrak, and they also provided preliminary observations on the internal anatomy of the female.
The material examined here all comes from Melinna elisabethae caught off the coast of northern Norway in Arctic latitudes from 67° to nearly 73° N and at depths of 74 to 371 m. This is a new host record for M. ergasiloides . The new material conforms closely to M. ergasiloides as redescribed by Bresciani & Lützen (1975) but there are some differences: Bresciani & Lützen (1975) showed the antennules of the female as elongate and indistinctly 6-segmented, quite similar to that of the male. The antennules of females from M. elisabethae examined here are much shorter and indistinctly 2-segmented. These reduced limbs are difficult to observe in the large females so we tentatively interpret this difference as due to the state of preservation of the material.
We also differ from Bresciani & Lützen (1975) in our interpretation of the oral area of the adult male, in particular, in the form of the mandibles. The oral region of the male includes a pair of small processes, referred to here as labral processes, which were clearly visible in Bresciani & Lützen’s illustration (1975, fig. 6) and were referred to as sinuous structures continuous with the anterior margin of the buccal armature. Bresciani & Lützen (1975) described the mandibles as unsegmented and terminating in a “broad comb and blunt anterior-directed projection”. We were unable to observe a comb, but the surface of the rounded basal part was densely ornamented with spinules in our material. The identification of this paired structure as mandibles is based on inferences made by Bresciani & Lützen (1975), who examined the transitional developmental stage, the copepodid.
The female in figure 14A carries three conical structures attached to the ventral body surface in the genital region (arrowed in Figure). Each has a circular base which adheres to the body surface of the female and a tapering distal part that may be slightly twisted. We identify these structures as spermatophores and, after studying the male of Lanassicola arcticus gen. et sp. nov. described below, we propose a novel method of deposition for spermatophores in male saccopsids (see below).
The female parasites on M. elisabethae were all attached laterally in the mid-body region around the junction between thorax and abdomen of the worm, in the vicinity of the 18th setiger, as illustrated by Bresciani & Lützen (1975: Fig. 1 View FIGURE 1 ) for their material of M. ergasiloides from Melinna cristata .

| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |