
Hawaiiandra puncticeps ( Sharp, 1878 ), 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.5164485 |
|
DOI |
https://doi.org/10.5281/zenodo.8400183 |
|
persistent identifier |
https://treatment.plazi.org/id/975887B7-FFEC-FFD7-66D0-FC38104F3096 |
|
treatment provided by |
Felipe |
|
scientific name |
Hawaiiandra puncticeps ( Sharp, 1878 ) |
| status |
|
Hawaiiandra puncticeps ( Sharp, 1878) View in CoL
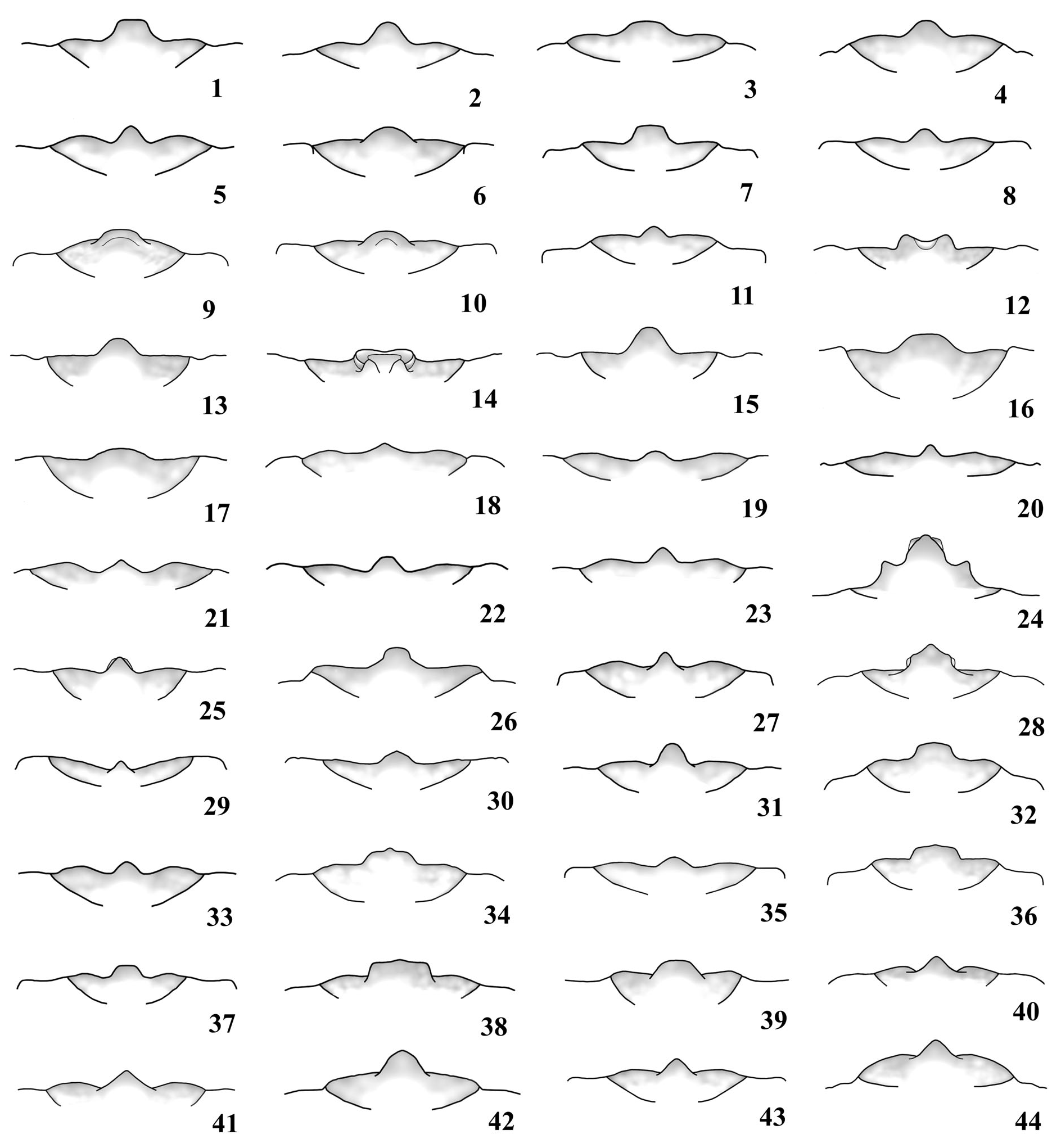
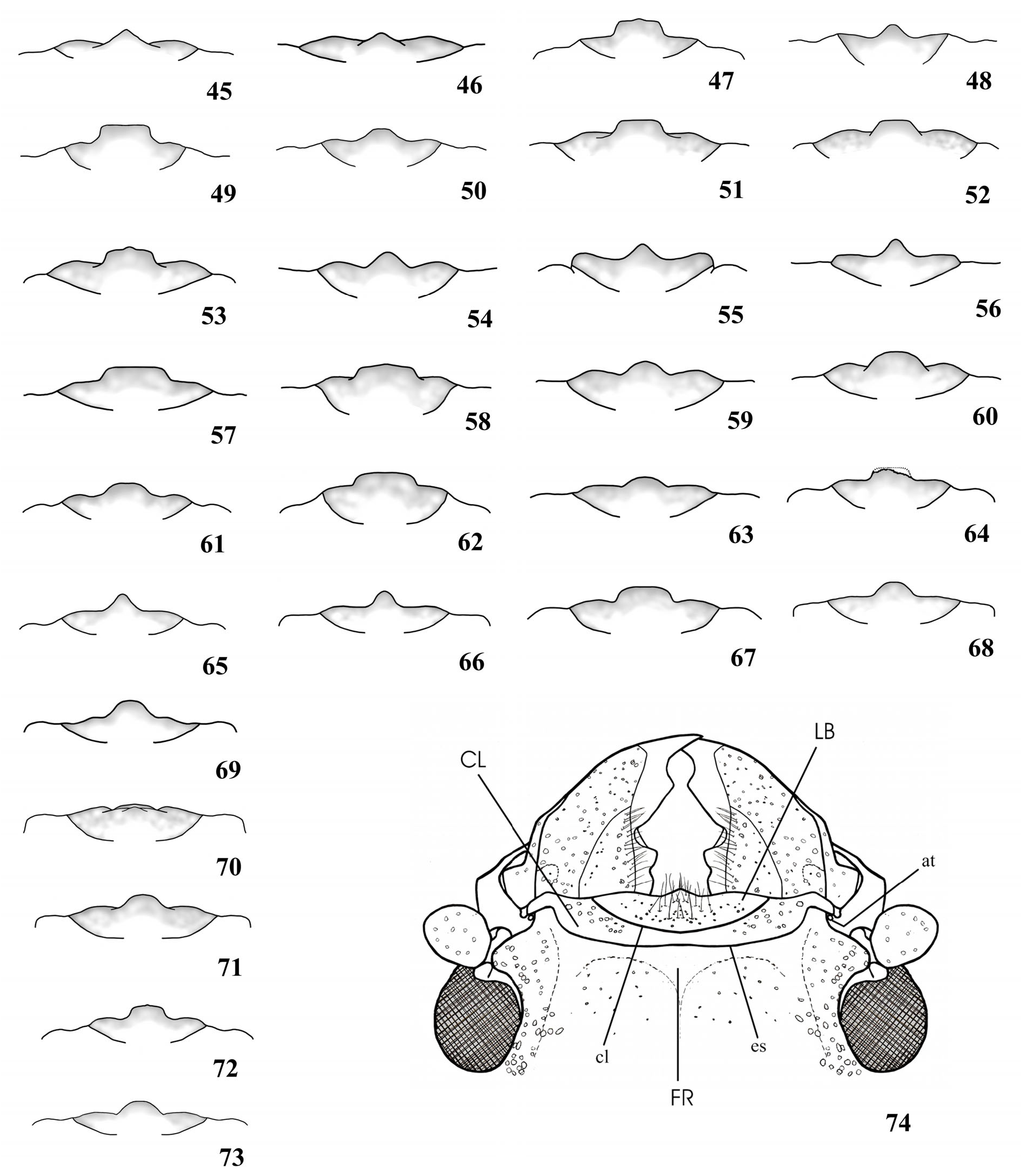
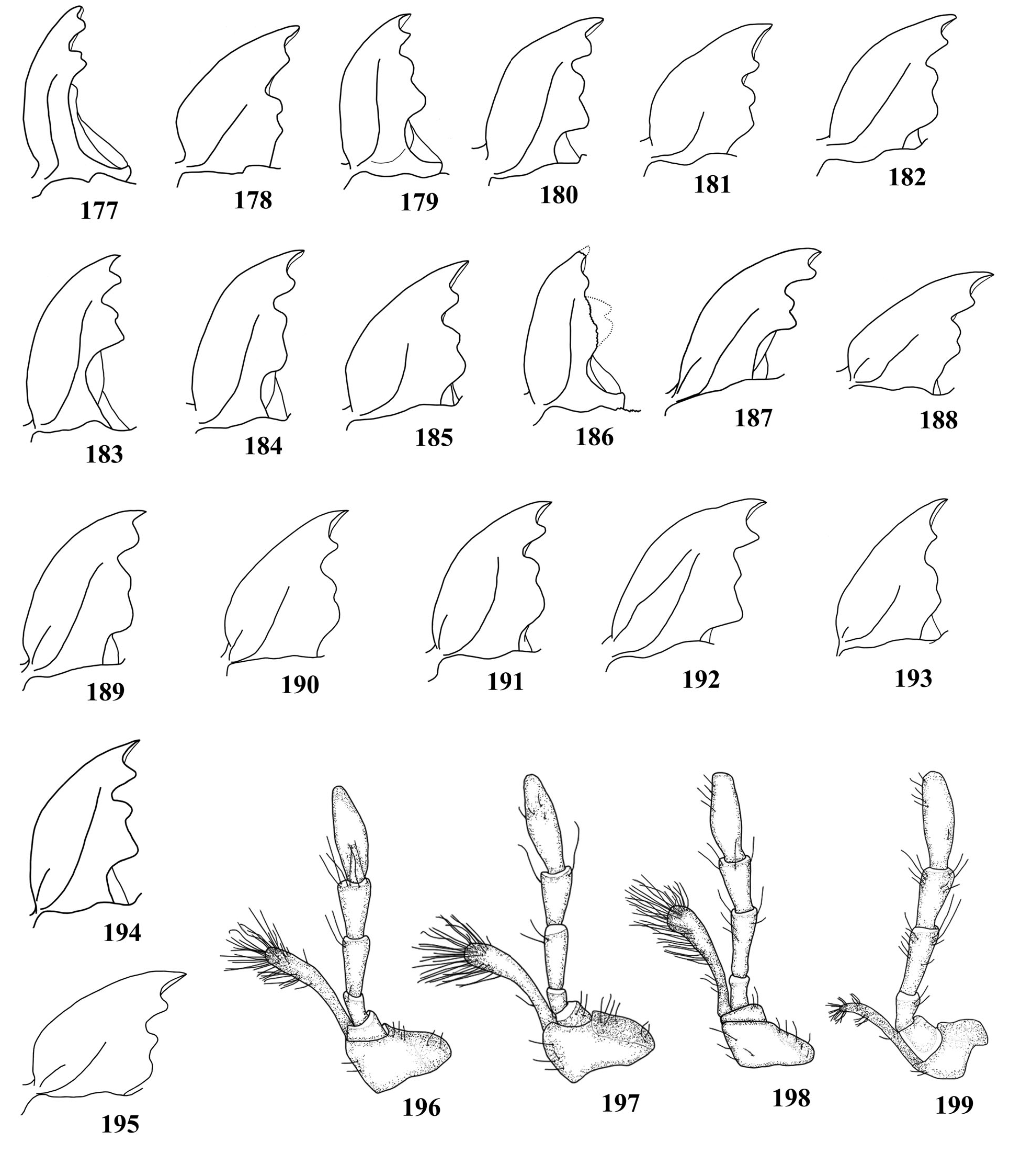
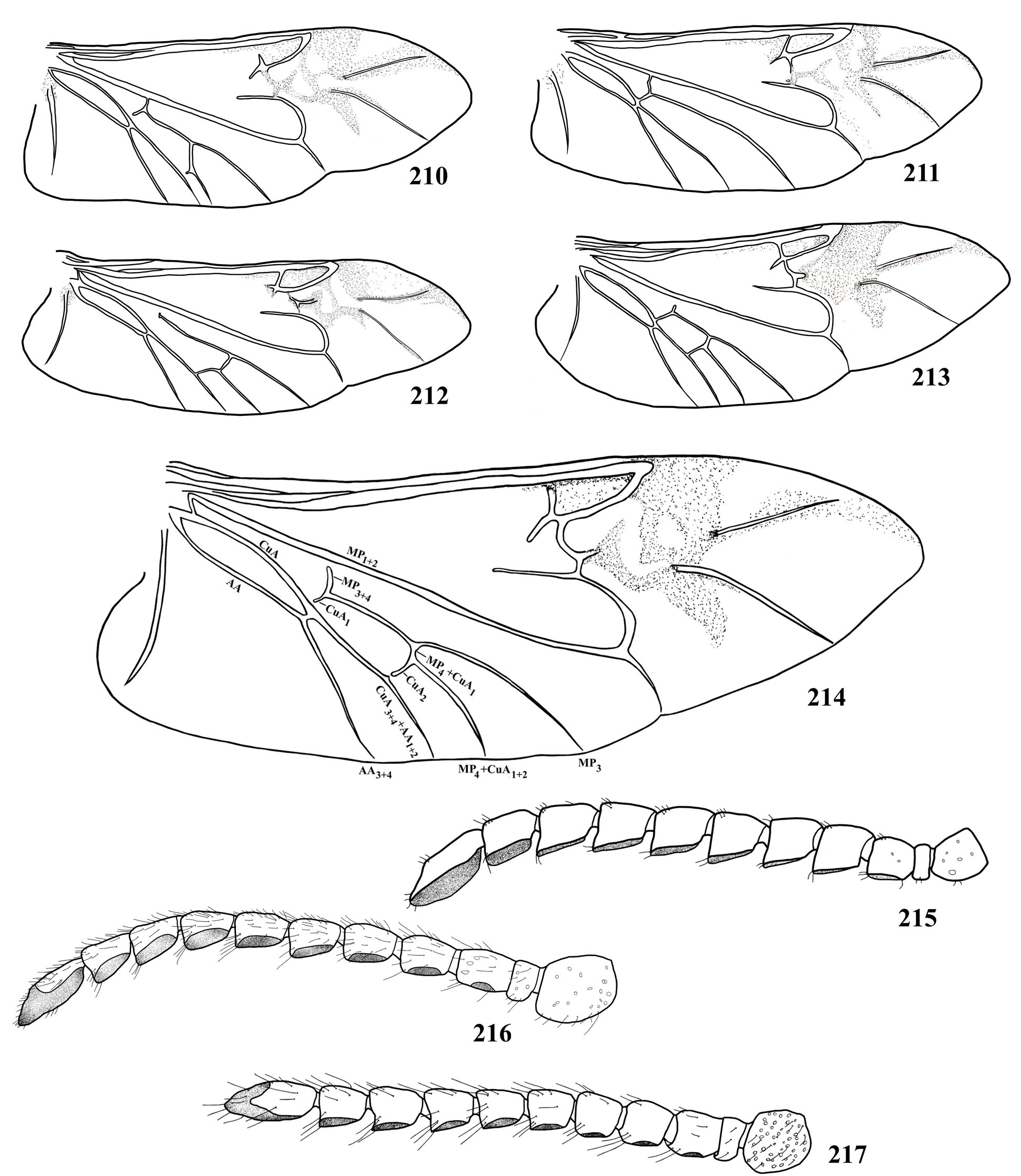
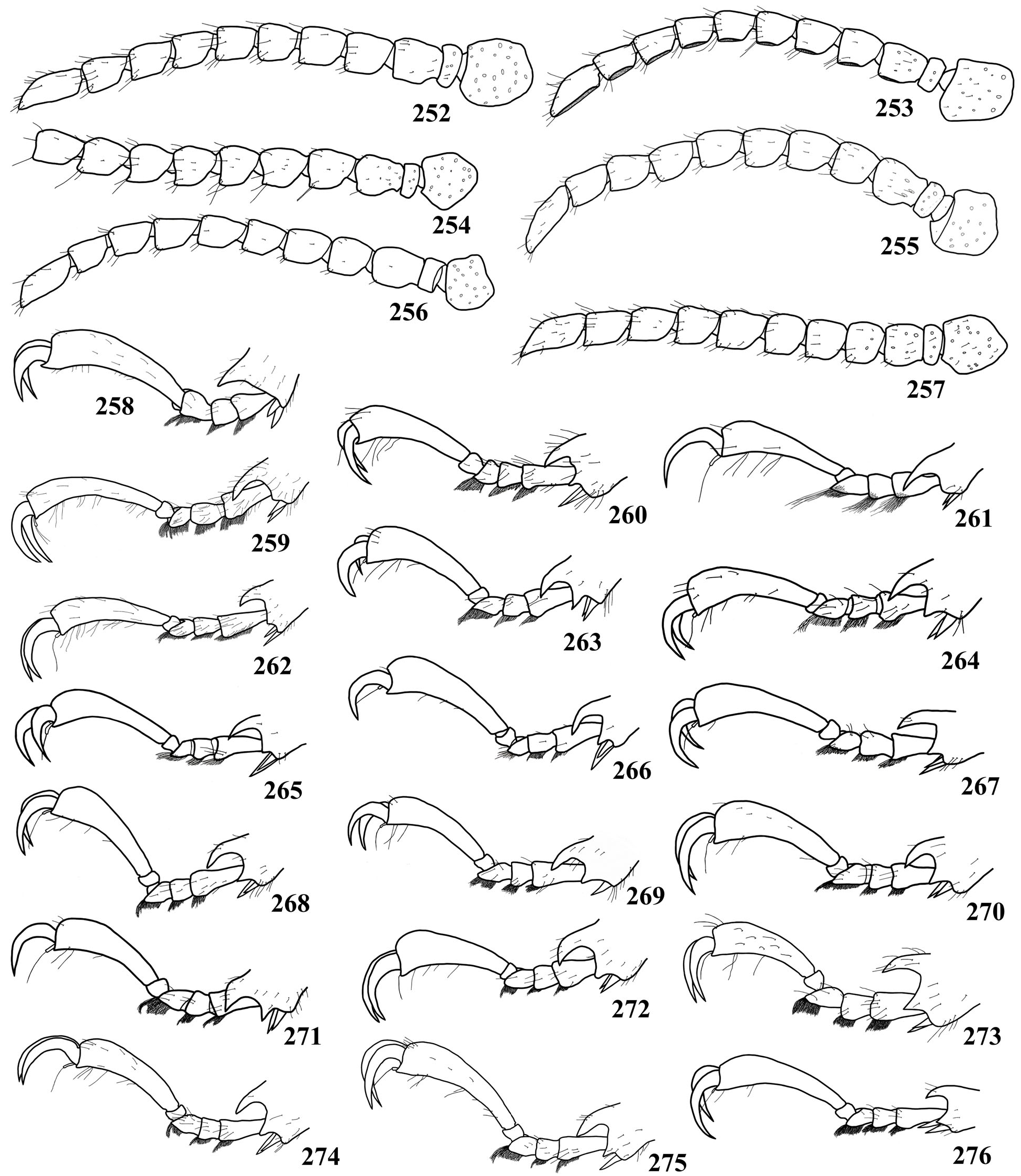
( Fig. 30 View Figure 1-44 , 74 View Figure 45-74. 45-73 , 75 View Figure 75-89 , 123 View Figure 118-147 , 199 View Figure 177-199. 177-195 , 209 View Figure 200-209. 200-204 , 216 View Figure 210-217. 210-214 , 259 View Figure 252-276. 252-257 , 345, 347-353)
Parandra puncticeps Sharp, 1878: 202 View in CoL ; Blackburn and Sharp 1885: 260; Sharp 1900: 95; Koebele 1901: 61; Lameere 1902: 82; Terry 1905: 73; Perkins 1907a: 96; 1912: 257; Lameere 1913: 5 (cat.); Giffard 1914: 14; Lameere 1919: 17; Swezey 1921: 170; Giffard 1922a: 28; 1922b: 118; Swezey 1922: 29; 1925: 198; 1931: 496, 498; 1935: 92; Ford 1952: 358; Duffy 1953: 141; Swezey 1954: 11, 60, 161, 217; Gressitt and Davis 1970: 388; Ibara 1972: 156; Gressitt and Davis 1972: 4; 1973: 214; Davis 1973: 157; Simon et al. 1984: 12; Suehiro 1986: 51; Stone and Pratt 1994: 281; Santos-Silva 2002: 32.
Birandra (Birandra) puncticeps View in CoL ; Santos-Silva and Shute 2009: 32.
Description. Integument shining, dark-brown; parts of head, mandibles, pronotal margins, elytral suture, apex and lower edge of femora and of tibiae blackish.
Male ( Fig. 347, 348 View Figure 347-353 ). Head proportionally large ( Fig. 348 View Figure 347-353 ) or very large ( Fig. 347 View Figure 347-353 ), almost as wide as apex of pronotum. Center of dorsal surface of head punctate: with punctures coarse and sparse, between clypeus and posterior edge of eyes; coarsely punctate, in part confluent, laterally, close to eyes; in central area, between posterior edge of eyes and occiput, with punctures barely coarse and more abundant than in area between the eyes. Dorsal face of head, close to basal edge of clypeus, with smooth transverse strip that expands triangularly at central area. Area behind eyes coarsely, abundantly punctate (punctures finer than in dorsal area close to eyes), in part confluent. Clypeus and labrum with long, moderately abundant hairs. Submentum depressed, coarsely and moderately sparse; anterior edge distinctly elevated; pilosity moderately long and abundant. Mandibles as long as head ( Fig. 123 View Figure 118-147 ). Eyes as in Fig. 75 View Figure 75-89 . Central projection of labrum ( Fig. 30 View Figure 1-44 ).
Pronotum 1.5 times wider than long; lateral margins sub-parallel at anterior two-thirds; central area of disc finely and sparsely punctate; lateral with punctures barely coarser and moderately abundant, mainly near posterior and anterior angles; anterior margin slightly concave at central region. Elytral punctation sparse. Metasternum with punctures slightly coarse, barely abundant close to metepisterna, finer and sparser towards the metasternal suture. Metepisterna coarsely, abundantly punctate at basal fourth, gradually finer and sparser towards apex. Metafemur ( Fig. 349 View Figure 347-353 ) short and enlarged; lower edge of femurs with moderately long, abundant hairs, mainly in metafemora. Tibiae clearly enlarged at apex. Metatarsus (without claws) longer than metatibiae; metatarsomere V ( Fig. 259 View Figure 252-276. 252-257 ) longer than I-III together.
Female ( Fig. 350 View Figure 347-353 ). Dorsal surface of head, pronotum, and elytra with same sculpture as male, and with same variations. Submentum depressed, coarsely punctate, abundant and confluent close to anterior edge, and more scattered towards gula; pilosity long and moderately abundant; anterior edge gradually elevated. Lateral margins of pronotum slightly rounded and convergent between lateral and anterior angles.
Variability. Male: center of dorsal surface of head, between clypeus and posterior edge of eyes, coarsely punctate, slightly sparser than in area behind eyes; clypeus with sparse hairs; mandibles from as long as head to longer than head; submentum not depressed and without elevation at anterior margin; punctures of submentum moderately abundant, in part confluent; pronotum from 1.4 to 1.6 times wider than long; lateral margins of anterior two-thirds of pronotum from convergent to divergent; punctures of central area of pronotum fine or very fine, sparse or abundant; pronotum laterally with moderately fine and sparse punctures, and barely coarse close to posterior and/or anterior angles; anterior margin of pronotum from slightly concave at central area to distinctly concave throughout; elytral punctation barely fine and, frequently, moderately abundant; metasternum finely and sparsely punctate throughout, including, close to metepisterna; tibiae not strongly enlarged at apex. Female: submentum with hairs moderately sparse.
Dimensions in mm (M / F). Total length (including mandibles), 22.0-26.5/26.0-30.3; prothorax: length, 4.4-5.0/5.3-5.8; anterior width, 5.5-7.2/6.2-7.7; posterior width, 5.1-6.3/6.9-7.9; humeral width, 5.9-7.5/ 8.0-9.3; elytral length, 13.1-14.4/16.3-18.5.
Geographical distribution. Hawaiian Islands (Hawaii, Kauai, Maui, Molokai, Oahu).
Material examined. (4 M, 4 F), as follows: USA, Hawaii: M, I.14.1929, ( MZSP) ; M, F (syntypes), [no date or name of collector] ( IRSN) ; Oahu Island : Mount Kaala, M, VII.5.1957 ( BPBM) ; Judd Trail, F, VIII.14.1919, O. H. Swezey coll. ( BPBM) ; Tantalus, F, IV.23.1905, W. M. Giffard coll. ( BPBM) ; 1 M, 1 F, V.14.1978, [no collector indicated] ( MTCO). Big Island : 29 miles Olaa ; 3800 ft; collected as pupa - reared until adult eclosion in VIII.1921 – specimen commented by Giffard 1922b), M, VII.1921, W. M. Giffard coll. ( BPBM) ; 29 miles Kilanea , F, II.10.1912, W. M. Giffard coll. ( BPBM) .
Types, type locality. We have no conclusive proof of where any of the specimens used in the original description are located. Evidence points to the specimens being in the BMNH, however it is possible that some may be in the IRSN or the MNHN. Sharp (1878) described Parandra puncticeps based on two males and, probably, a single female: “In the female the length of this segment [apical ventral segment] is considerably greater, and its hind margin is obscurely truncate in the middle; the punctures are less obsolete, and the abundant ciliae of the hind margin are twice as long as in the male. Of this latter sex I have two individuals before me, one small the other large”. All specimens are from Hawaii, Oahu Island .
We received a couple of “ syntypes ” from IRSN, from Hawaii (without precise locality), and one of the authors ( Matsuda ) has examined a “ syntype ” female in MNHN, from Oahu Island. However, we suspect that the true syntypes of P. puncticeps are deposited at BMNH ( Fig. 351-353 View Figure 347-353 ). Sharon Shute ( BMNH, pers. comm.) wrote: “I have found three specimens of puncticeps Sharp that are definitely syntypes. There is one male double mounted on the original card, and on the card is written “male a. Ins Oahu Blackburn 1877, Type D.S.” ; there is an additional BM printed label “ Hawaiian Islands , Blackburn”, and a BM registration number “Sharp 1905-313”. There is a female with the same data plus a small male where the card has been cut and put below the specimen ; on the card it states “male b. same data” .
It is important to point out that the calligraphy on labels of the specimens mentioned by Sharon Shute, agrees very well to that figured by Horn and Kahle (1936) of a label by David Sharp.
Horn and Kahle (1936) wrote on Sharp’s collection: “Sharp, David (1840 – 1922), Lamellicorn. der I. Sammlg. an R. Oberthür (Rennes). – Cerambycid. ex parte via E. Le Moult, via P. Boppe zurück an E. Le Moult (Paris); ex parte via Janson & Sons, via J. Archad, via P. Boppe an E. Le Moult (Paris). – Spezial-Sammlg. brit. Coleopt. via Tochter an Brit. Mus. N. H., London. – Rest 1907 an Brit. Mus. N. H., London. – Hinterlassene Privat-Sammlg. Curculionid. der Welt ex parte an Brit. Mus. N. H., London; ex parte 1927/28 an Exper. Station. Hawaiian Sugar Planters Assoc., Honolulu”.
In spite of the statements above, Cambefort (2007) has not recorded the Boppe Collection or the Le Moult Collection in MNHN. However, there are some specimens from the Le Moult Collection deposited at MNHN, but usually in an indirect manner explained by Cambefort (2007).
On the Sharp Collection, Cambefort (2007) wrote: “As collection de coléoptères, très riche em exemplaires des colonies anglaises, fut dispersée. Son ami Oberthür put acquérir le “premier choix” des scarabéides, avec les nombreux types qu’ils refermaient”. That information agrees with that by Horn and Kahle (1936) about the lamellicorns of Sharp’s Collection.
Based on the personal communication of Sharon Shute and pictures of the specimens deposited at BMNH, we can only presume that the “ syntypes ”, deposited at IRSN, are not true types. Unfortunately, we can only suppose that the female “ syntypes ” deposited at IRSN and MNHN are not true types. It is important to mention that the calligraphy on the labels of the specimens deposited at IRSN does not agree with that on a Sharp’s label figured on Horn and Kahle (1936: plate XXIV, fig. 36).
The “ syntype ” female deposited at MNHN, although being a female of P. puncticeps , was glued. It is evident there is glue joining the prothorax with the remaining body. Besides, the head is not of P. puncticeps , and not of a female: it is a head of a male of Komiyandra , probably K. javana .
From above, we believe that the true syntypes of Parandra puncticeps are deposited at BMNH, but we are not designating a lectotype at this time because we are unable to absolutely prove that the specimens are really the syntypes (one or all).
Comments. Sharp (1900), was the first to write on the extreme variation: “This species exhibits a great deal of variation in the form and proportions of the prothorax, epistome, mandibles, etc. and there may possibly be more than one form in the islands. The material before me is not sufficient to decide as to this, owing to the development of the individual being subject to much variation; but the varieties appear to be to some extent located in different islands”.
Later, Giffard (1922b) who collected and reared some specimens at Oahu, wrote: “Although the mandibles and the lateral margins of the thorax are extremely variable in male examples from each island, there are intermediate forms which connect these extremes. This is quite noticeable both as to structure and sculpture in the eighteen reared specimens from Hawaii previously referred to. The smaller series taken “in situ” on Oahu and Kauai present the same tendencies. The representative collections of this Cerambycid have heretofore been very sparse in individual specimens, and in consequence many of the variations noticed from time to time have led some to suspect the possibility of more than the one species described. Examination and study of a series like the present one, however, tends to lessen any such suspicion unless some other important but constant character than is yet known can be found by further study of larger series from all the islands in the archipelago”.
Contrarily, Gressitt (1978) recorded: “There are 2 other lines of endemic Hawaiian Cerambycidae , but these each represent a single species (each in a different subfamily: Parandrinae and Prioninae ) which have not proliferated at all. Both of these, Parandra puncticeps Sharp and Megopis reflexa (Karsch) , are found on all the major islands of the chain, and individuals from the different islands do not seem to have developed any local population characteristics”.
As in some species of Parandra , the form of the head and of the prothorax suffers from great variation in H. puncticeps , as pointed out by Sharp (1900). Those variations are not related with the geographical distribution, and so, they are only specific variation, that can be confirmed through the intermediate forms between the extremes. Although this shows that Sharp (1900) was wrong on the possible existence of different forms among the Hawaiian Islands, it also shows that Gressitt (1978) omitted that possibility or, at least, did not comment on the great specific variation in the species.
Also, without commenting on anything about the shape variation, Zimmerman (1942) wrote on Parandra puncticeps : “On the other hand, there are genera which have not developed species complexes in the islands. Parandra , for example, is considered one of the most primitive genera of the Cerambycidae (longhorn bettles [sic]) and is world wide in distribution. It has only one endemic species in Hawaii and only one in Fiji. I see no reason for considering that the ancestors of the Hawaiian Parandra arrived in Hawaii at any later date than did the ancestors of the Plagithmysus group; yet the Plagithmysus group has split up into six groups called genera and no less than 95 species. I do not know why Parandra should not have followed the course of Plagithmysus - but there is probably a good reason, hidden at the moment, which some geneticist might interpret in the future”. This comment was followed by Gressitt (1971): “The 122 or more endemic cerambycids of Hawaii have evolved from only three original ancestral lines. Two of these, representing the Parandrinae and Prioninae , have only one species each, whereas the third, in the Cerambycinae , has evolved extensively, adaptively radiating so that the species were assigned to eight genera”.
Curiously, nobody commented on the similar appearance of H. puncticeps , Archandra caspia and the species of Parandra , that share many characters that do not occur in the other genera of Parandrini , like the paronychium with more than one setae, shape of the mandible in females, and procoxal cavities almost closed (closed in Archandra and Parandra ).
Blackburn and Sharp (1885) commented on the possibility of H. puncticeps occurring outside of the Hawaiian Islands: “This species or a closely allied one occurs in the Philippines islands”. Although we do not know any species similar to H. puncticeps from the Philippines, there are two possibilities to explain the specimen(s) studied by Blackburn and Sharp (1885): specimen(s) wrongly labeled; or an evident error of identification. We do not know any work in which the Philippines has been formally included as an area of occurrence of H. puncticeps .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hawaiiandra puncticeps ( Sharp, 1878 )
| Santos-Silva, Antonio, Heffern, Daniel & Matsuda, Kiyoshi 2010 |
Birandra (Birandra) puncticeps
| Santos-Silva, A. & S. Shute 2009: 32 |
Parandra puncticeps
| Santos-Silva, A. 2002: 32 |
| Stone, C. P. & L. W. Pratt 1994: 281 |
| Suehiro, A. 1986: 51 |
| Simon, C. M. & W. C. Gagne & F. C. Howarth & F. J. Radovsky 1984: 12 |
| Gressitt, J. L. & C. J. Davis 1973: 214 |
| Davis, C. J. 1973: 157 |
| Ibara, W. 1972: 156 |
| Gressitt, J. L. & C. J. Davis 1972: 4 |
| Gressitt, J. L. & C. J. Davis 1970: 388 |
| Swezey, O. H. 1954: 11 |
| Duffy, E. A. J. 1953: 141 |
| Ford, E. J., Jr. 1952: 358 |
| Swezey, O. H. 1931: 496 |
| Swezey, O. H. 1925: 198 |
| Giffard, W. M. 1922: 28 |
| Giffard, W. M. 1922: 118 |
| Swezey, O. H. 1922: 29 |
| Swezey, O. H. 1921: 170 |
| Lameere, A. A. 1919: 17 |
| Giffard, W. M. 1914: 14 |
| Lameere, A. A. 1913: 5 |
| Perkins, R. C. L. 1912: 257 |
| Perkins, R. C. L. 1907: 96 |
| Terry, F. W. 1905: 73 |
| Lameere, A. A. 1902: 82 |
| Koebele, A. 1901: 61 |
| Sharp, D. 1900: 95 |
| Blackburn, T. & D. Sharp 1885: 260 |
| Sharp, D. 1878: 202 |