
Thylogale billardierii ( Desmarest, 1822 )
|
publication ID |
https://doi.org/ 10.1093/mspecies/sey012 |
|
publication LSID |
lsid:zoobank.org:pub:022B7817-9037-4B26-96FF-2D2488D63FB7 |
|
DOI |
https://doi.org/10.5281/zenodo.4593293 |
|
persistent identifier |
https://treatment.plazi.org/id/97546218-FFB1-FF8D-6DAC-CD0CFAE3FDC1 |
|
treatment provided by |
Felipe |
|
scientific name |
Thylogale billardierii ( Desmarest, 1822 ) |
| status |
|
Thylogale billardierii ( Desmarest, 1822) View in CoL
Tasmanian Pademelon
kangurus Billardierii Desmarest, 1822:542. Type locality “La terre de Van-Diemen” (= Tasmania).
Halmaturus (Thylogale) tasmanei Gray, 1838:108 . Type locality “Van Diemen’s Land” (= Tasmania).
Macropus (Halmaturus) rufiventer Ogilby, 1838:220 . Type locality “Van Diemen’s Land” (= Tasmania).
H [almaturus] brachytarsus Wagner, 1842:104, 121. Type locality unknown.
CONTEXT AND CONTENT. Context as above. No subspecies are recognized ( Calaby and Richardson 1988).
DIAGNOSIS
upper lip. The snout is naked and the tapering tail is covered with short hairs. The nasal bones of the skull have straight outer edges, each bone being wedge shaped ( Fig. 2 View Fig ). Females carry a single young in a forward-opening abdominal pouch with 4 teats; females are smaller but otherwise indistinguishable from males.
Driessen (1992), who used pouch young to determine maturity in females and sperm in the cauda epididymides maturity in males, reported mean weight of adult females to be 4.56 kg (n = 338; range 2.2–7.4 kg) and that of males to be 6.53 kg (n = 346; range 3.3–11.0 kg). From this large sample collected by shooters (Driessen 1992), we can conclude that T. billardierii has a 1:1 sex ratio.
Means and ranges of external measurements (mm) of male and female T. billardierii , respectively (n = 40, 33) were as follows: total length, 852.7 (720–1,150), 750.0 (720–870); length of tail, 361.4 (300–445), 312.5 (270–325); length of hind foot, 133.8 (110–160), 119.4 (97–145); length of ear, 63.5 (58–72), 58.8 (45–68).
Mean skull measurements (mm) for T. billardierii , male and female, respectively (n = 18, 15) were as follows: greatest length of skull, 110.2, 101.6; depth of rostrum, 20.1, 17.7; cranial breadth,15.9, 14.6; zygomatic breadth, 56.5, 50.6; length of maxillary toothrow, 29.6, 27.5; length of molariform teeth in mandibular toothrow, 27.1, 25.7.
In comparison to other macropodids, members of Thylogale are of medium size, usually 3–9 kg, with relatively large forelimbs compared with larger macropodids, long and fine fur, and a well-haired tail (with short stiff hairs) that lies on the ground when at rest ( Fig. 1 View Fig ). Although differing in coloration and distribution (see “Key”), all species are similar in size, with head and body lengths about 600 ± 100 mm SE (except the smaller T. calabyi —Calaby’s pademelon), tails about two-thirds the length of head and body, and same-age males larger than females. T. billardierii is the largest species in the genus (e.g., mean body weights of males:females of 9.0: 5.8 kg, those of T. stigmatica (red-legged pademelon) are 6.5: 4.2 kg, and those of T. thetis (red-necked pademelon) are 7: <4 kg—Menkhorst and Knight 2001) and is distinguished from other pademelons by its relatively shorter tail, its orange-to-rufous-red venter, and the absence of both facial and hip markings.
GENERAL CHARACTERS
Thylogale billardierii has a compact body form, short ears, and long pelage, which is dark-brownish gray above and yellowish or rufous below ( Wood Jones 1924). Facial markings are absent except for a faint yellowish line which runs along the
DISTRIBUTION
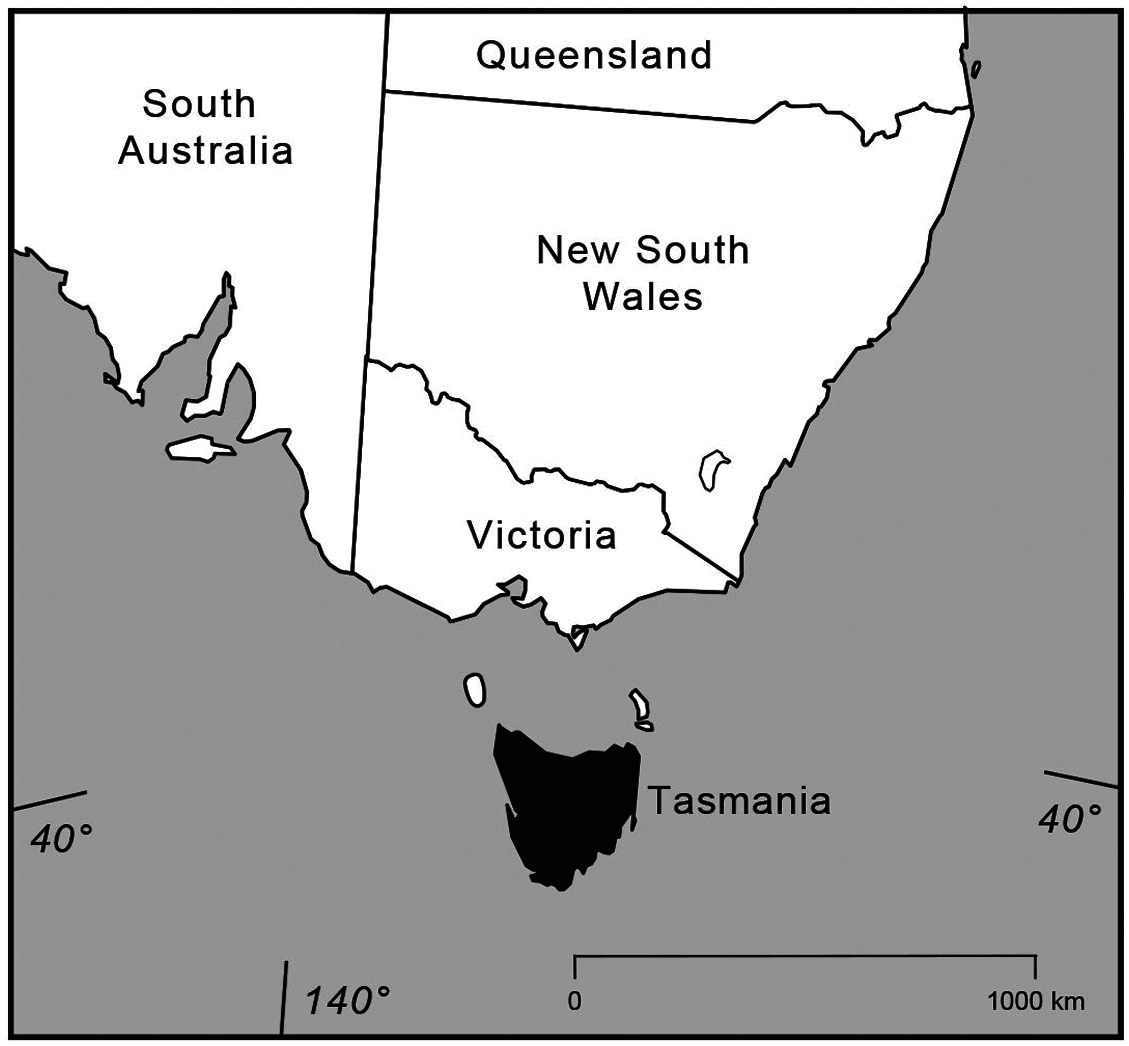
Extinct on mainland Australia for> 100 years, Thylogale billardierii is widespread and often is exceedingly common in Tasmania ( Fig. 3 View Fig ). It is found in a greater percentage of 100- km 2 blocks in the state than all but one other species (tied with Macropus rufogriseus , red-necked wallaby, at 41%— Rounsevell et al. 1991). Populations are present on the King Island cluster off the northwest coast and on Flinders, Cape Barren, Clarke, Prime Seal, East Sister, and West Sister islands in the Furneaux Island group off the northeast coast ( Hope 1973; Johnson et al. 1989). Spotlight (road) surveys indicate that T. billardierii increased in abundance between 1975 and 1990 in Tasmania ( Driessen and Hocking 1992). Thus, although populations from southeastern South Australia and Victoria have been extirpated, T. billardierii seemingly was secure in Tasmania until the intentional introduction of Vulpes vulpes , the red fox (by one or a few individuals), into the state in 2001 or 2002.
FOSSIL RECORD
The ancestry of the genus Thylogale can be traced to the early Pliocene ( Flannery 1984), together with other genera surviving today in wet or well-watered regions. The formerly limited distribution of Thylogale billardierii in coastal Victoria strongly suggests that the species arose in Tasmania and invaded the mainland during a period of lower sea level ( Maynes 1989). Fossil pademelons of late Pleistocene age (30,420 –11,630 years ago) have been recovered from Nunamira Cave in south central Tasmania ( Cosgrove et al. 1990), the species is known from both Pleistocene and recent locations in McEachern’s Cave in extreme southwestern Victoria ( Wakefield 1967), and Wakefield (1964) reports subfossils from 3 of 7 sites on the basalt plains of western Victoria, including both cave and aboriginal (beach midden) sites. One of the latter yielded an estimated 44 individuals.
FORM AND FUNCTION
The body of Thylogale billardierii is squat, the tail is thick and relatively short, and its fur is dense. Narrow eye rings are tinged with rufous, and the underparts are buff to orange, including mouth and chin, with rufous tinge, especially on the belly.
The dental formula is i 3/1, c 1/0, p 1/1, m 4/4, total 30. The dentition is typical of the Diprotodontia , in which the lower incisors are reduced to a single pair of forward-projecting teeth. All teeth have roots, and hence, animals can be aged by degree of tooth wear. A more detailed method of aging by molar tooth eruption of known-aged live animals proved useful for ages 5–30 months, by which time the 4th molar is erupting ( Driessen and Hocking 1997).
Thylogale billardierii is a low-fiber browser that also includes some grass in the diet ( Sanson 1989). The incisors can clip certain plant parts, but highly lignified petioles of leaves of dicotyledonous plants require shearing with the cheek teeth. The small premolars and the molars (both with shearing and grinding capacities) lie in a flat toothrow in which all teeth occlude at once ( Sanson 1989).
The long gastrointestinal tract is typical of foregut-fermenting macropodines, but the sacciform forestomach is not as massive as those of other Thylogale species ( Dellow 1979). T. billardierii also differs from its congeners by possessing a gastric sulcus, a feature that may function to channel ingested milk directly to the hindstomach in young animals ( Dellow 1979).
The field metabolic rate, using the doubly labeled water method, was 0.532 ml CO 2 g−1 h−1, or 1,630 kJ/day ( Nagy et al. 1990), or an average of 273 kJ kg−1 day−1 (B. Green 1989). Water influx rates estimated to be 108 ml kg−1 day−1 were required to maintain the 70.3% water content of the body. Nagy et al. (1990) estimated the dry matter digestibility of grass to be 56 ± 5% (SE), and that a 5.9-kg wild T. billardierii would have to consume 196 g of dry matter of food each day to meet its metabolic energy requirements.
The use of indwelling jugular venous catheters confirmed that cortisol is quantitatively the major circulating glucocorticoid, with the mean concentration being high (200–250 nmol/ ml) in undisturbed animals (Martin and McDonald 1986). Glucocorticoids affect carbohydrate metabolism by both antagonizing the peripheral actions of insulin and enhancing hepatic gluconeogenesis by induction of gluconeogenic enzymes ( Martin and McDonald 1988). Thus, the normal role of glucocorticoids may be to permit metabolic processes to achieve maximum rates and to synergize other hormones to achieve metabolic control ( Martin and McDonald 1988). Maximal adrenocorticotropic hormone stimulation via intravenous infusion briefly caused a 5- to 6-fold increase in cortisol secretion rate, but this dropped by one-half due to a marked increase in metabolic clearance. Plasma glucose concentration did not change significantly because of infusion of either adrenocorticotropic hormone or plasma cortisol (Martin and McDonald 1986, 1988).
ONTOGENY AND REPRODUCTION
Parturition in Thylogale billardierii is typical of macropodids; the mother’s tail is extended forward while she licks her urogenital sinus and pouch. Once the 420-mg neonate (crownrump length of 1.62 cm) reaches the pouch, it attaches to 1 of the 4 teats, where it begins a long pouch life of nearly 7 months. Teat selection is random except that the large teat of a previous lactation is avoided ( Rose and McCartney 1982b).
As happens with other marsupials, the milk of T. billardierii changes ( Rose and Flowers 2005) from dilute milk in early lactation (0–6 weeks: 500–800 kJ/ml) to more concentrated milk in midlactation (17–25 weeks: 550–1,100 kJ/ml) and more energy rich in late lactation (≥ 26 weeks: 1,300 –2,550 kJ/ml). The amounts of carbohydrates, similar in early lactation and midlactation, drop in late lactation, whereas those of proteins and lipids rise significantly in late lactation compared with earlier periods. Milk carbohydrates were always higher in the captive group compared with T. billardierii from several localities in the wild. Although protein and lipid levels were more variable in the captive group, the energy content of the milk led to faster growth rates than those seen in the wild ( Rose and Flowers 2005). The premature pouch vacation in females receiving bromocriptine injections (near the end of the pouch life of her young) indicates a maternal role in final pouch vacation ( Rose 1985, 1997).
The sex of young can be determined by day 14 with the unaided eye, using the criterion of pouch anlage or scrotum. Although the pigmented eyeballs can be seen at birth, the eyelids and conjunctiva are not evident until week 5, the follicles of the eyelashes are seen at week 9, and the eyelids do not open until between weeks 18 and 20. The pinnae are free from the underlying skin by the 5th week but are not fully uncurled until 18–20 weeks ( Rose and McCartney 1982b).
Other aspects of development are slow too, such as the lower incisors 1st appearing by day 91 and the 1st squeaking vocalizations at day 96. Pigmentation appears on head, ears, and tip of tail between days 60 and 115, and by day 140, the entire dorsum is pigmented. Between days 144 and 160, the body becomes covered with fine fur, after which a top coat of guard hairs appears. Shortly after becoming well furred (usually 150–160 days), the young’s head protrudes from the pouch on a regular basis. Although sufficiently coordinated to be able to stand unaided before this age, about this time the young is able to hop. The minimum and maximum ages for first emergence from the pouch were 158 and 192 days. After vacating the pouch, the young remains at foot for another 3 months, continuing to suckle during this period ( Rose and McCartney 1982b).
The relative growth rate (2.37 × 10 −2 log e g/day) is typical of macropodids and enables the young to permanently exit the pouch by 202 ± 7 days (range 196–212 days). At pouch vacation, the average young weighs 1,029 ± 112 g SE (range 857– 1,170 g—Rose and McCartney 1982b). The young is weaned at 240–300 days, at a weight of about 3 kg. After permanent pouch vacation and the birth of the next neonate, the female has a pouch young attached to 1 teat and a young at foot nursing on a centimeter-long teat. Thus, although a female devotes nearly a year to raising an offspring, by that time she also has a pouch young nearly 4 months old. All things going well, a female raises about 1.3 young per year. Young born in winter survive better than those born in spring or summer (Driessen 1992). Animals continue to grow, if slightly, throughout life, enabling males to attain 11 kg ( Driessen and Hocking 1997). Life span in the wild is unknown.
It is unclear whether T. billardierii from Victoria might have been physically larger than those surviving on Tasmania. However, fossils of specimens from Victoria support that viewpoint ( Hope 1973). The smallest animals are from the smallest islands of the Furneaux Island group in the Bass Strait.
In Tasmania, T. billardierii can be hunted except during the few weeks of the deer season. Driessen (1992) reports that the loss of large reproductively mature adults in hunted populations was largely compensated by the earlier attainment of maturity than in populations not hunted. Thus, hunting altered the age distribution but not the level of breeding or, probably, population density.
The majority of young T. billardierii are born in autumn, although both in nature and in captivity some young are born in other seasons ( Rose and McCartney 1982a). A minor peak is seen in spring when young are leaving the pouch and are being either replaced by recently activated blastocysts or by young from new matings. Driessen (1992) recorded births in all months (n = 338) but with 2 distinct periods, April–June and October– November. During autumn and winter, first-time breeders give birth a month later than experienced breeders that failed to raise a young the previous breeding season. This can be explained by the median age of maturity for females being 13 months. In contrast, during the October–November peak, the mean birth dates did not differ between the 2 groups of females. The 2nd peak is probably due, in part, to the activation and subsequent birth of embryos produced at postpartum matings in autumn.
The anatomy of the male reproductive system is similar to that of other macropodids, with a pendulous scrotum lying anterior to the opening of the sheath housing the penis ( McCartney 1978). A large carrot-shaped prostate gland, a bipartite bulbourethral gland, and 3 pairs of Cowper’s glands are the accessory glands. The prostate gland increases in size in association with the autumnal breeding peak and to a lesser extent in the minor spring peak. The penis ends in a blunt cone and is retracted into a preputial sac.
Males are fertile soon after spermatorrhea, usually at 4 kg in males 14 or more months old ( Rose and McCartney 1982a). Driessen (1992), using tooth eruption as a criterion for aging, determined that the median age to maturity of 346 males was 17 months, with only 3% being mature at 12 months. Once sexually mature, males are continuously spermatogenic ( Setchell 1977).
The mature spermatozoan has a mean length of 98.8 µm, with a maximum head length of 7.25 µm ( Horak 1980). The sperm head is unusually shaped, being bluntly pointed in dorsal aspect, broad in the mid-region, and tapering posteriorly. The midpiece is convex, in part due to the ventral insertion of the flagellum ( Horak 1980).
Although the median age of maturity is 13 months for females (Driessen 1992), about 30% are mature at 12 months. Drought delays maturity in both sexes. In captivity, females as young as 14 months and as light as 3.5 kg gave birth, but most females were older and heavier when bearing their 1st offspring ( Rose and McCartney 1982a). Of 9 females evaluated for reproductive activity for 3 years while in captivity, some were in estrus in every month, mating occurred in all by 1 month, and births were recorded in March, October, and November ( Rose and McCartney 1982a).
The structure and shape of the female reproductive tract are described by Horak (1980) and McCartney (1978) and the origin, distribution, and structure of related blood vessels by Lee and O’Shea (1977). Four mammae are located 2 on each side in the pouch.
Ovulation of a single ovum is spontaneous, with a mean ovulation rate of 30.3 days in adults ( Rose and McCartney 1982a). The zygote develops to the blastocyst stage, after which embryonic growth proceeds by embryotrophic nutrition until a brief implantation during the last one-third of the 30.2-day gestation. Pouch life is about 7 months, and a young at foot is nursed for a further period ( Rose and McCartney 1982b).
Normal gestation occupies all of the follicular and luteal phases and is followed by a postpartum estrus and ovulation ( Rose and McCartney 1982a). If lactation follows, the corpus albicans associated with pregnancy declines in the same way as in the nonpregnant female, but the new corpus luteum formed at the postpartum ovulation is held in a state of quiescence, which may persist for almost the entire period of pouch occupancy (mean: 202 days).
Activation of the diapausing blastocyst is stimulated by reduced suckling, and thus, birth occurs the same night as final pouch vacation. Birth is always followed within 24 h by a postpartum estrus, so that parturition and mating usually occur during the same night ( Rose and McCartney 1982a). The resulting zygote develops into a 70- to 80-celled vesicular unilaminar blastocyst of about 280 µm diameter, which then enters embryonic diapause ( Horak 1980) until the young vacates the pouch. If the single pouch young is removed or lost, the next young is born about 27–28 days later ( Rose et al. 1999).
ECOLOGY
Despite going extinct on the mainland during the 19th century ( Calaby 1971), Thylogale billardierii has the broadest distribution of any mammal within Tasmania ( Rounsevell et al. 1991), suggesting wide habitat tolerances. However, it is most abundant where open lands (such as sheep farms) are mixed with forest lands. At night, T. billardierii moves from daytime refuges in dense cover to feed in adjoining open areas, including crop fields and pastures.
Thylogale billardierii is a generalist herbivore, grazing and browsing. The succulent short grasses and forbs and other foods (fruits, ferns, leaves, and twigs) all are considered to be low in digestible fiber. Sprent and McArthur (2002), who analyzed forestomach contents of animals feeding naturally near a young pine plantation, reported that, as a percentage of dry matter, T. billardierii ate 53% grasses and 38% broad-leafed forbs, 0% trees, and small amounts of other foods. However, T. billardierii selectively ate forbs because they were twice as numerous in the diet as in the field. In feeding trials of leaves and stems from seedlings of several tree species, individuals generally selected Acacia melanoxylon and Pinus radiata stems and leaves over Eucalyptus , but their browsing reduced the height of eucalypt seedlings by 17% compared with seedlings in exclosures ( McArthur et al. 2000). T. billardierii is a significant source of mortality and reduced growth in forest plantations of eucalypts.
Food regurgitation (merycism), a common behavior in macropodines apparently related to their foregut fermentation, is practiced by T. billardierii ( Mollison 1960) . Usually the food spilled from the mouth is reingested immediately, but Clancy (1982) observed 2 instances in which food regurgitated by females was eaten by 2 males and 1 female.
Thylogale billardierii can reach densities of about 8 individuals/ha, with overlapping home ranges of 29.7 ha for males and 14.8 ha for females ( leMar and McArthur 2003). Thus, besides a state-wide distribution, they are also abundant. In the middle one-third of the 20th century, millions of T. billardierii pelts were shipped to the northern hemisphere for use in the fur trade. T. billardierii meat also is eaten.
Thylogale billardierii is preyed upon by feral cats ( Felis catus — Fancourt 2015) and dogs ( Canis lupus familiaris ), Tasmanian devils ( Sarcophilus harrisii ), and formerly by thylacines ( Thylacinus cynocephalus ) in Tasmania and by red foxes and dingoes ( Canis lupus dingo ) on the mainland. Young T. billardierii are taken by spotted-tailed quolls ( Dasyurus maculatus ) or eagles too. Another source of mortality in Tasmania is the pesticide “1080” (sodium fluoroacetate, which inhibits citrate and succinate metabolism in the Krebs’ cycle, causing cardiac failure), which is used to control marsupial herbivores where eucalypt seedlings are planted. Such programs are effective, resulting in population declines of 97–98% following a 17-day baiting program ( leMar and McArthur 2001). Carcasses of 8 of 10 radiocollared T. billardierii killed during a 1080-poisoning operation were recovered an average of 31 m from the bait line ( leMar and McArthur 2000).
Likely competitors include red-necked wallaby, brushtail possum ( Trichosurus vulpecula ), and to lesser extents wombat ( Vombatus ursinus ) and introduced European rabbit ( Oryctolagus cuniculus ); all have been studied extensively as potential sources of mortality for young trees ( McArthur et al. 2000).
An unidentified species of large strongylate nematode in the genus Labiostrongylus causes lesions in the stomach of T. billardierii ( Munday 1971) . Larval stages of Baylisascaris tasmaniensis , an ascaridoid nematode, have been found in tissues of T. billardierii ( Spratt et al. 1991) . Seven species of ectoparasitic mites of 4 genera and 3 families were detected by R. H. Green (1989), namely, Cytostethum inerme , C. maydenenses , C. postsqumatum , and C. thylogale , Guntheria kallipygos , Neotrombicula novaehollandiae , and Thadeua mitchelli . Speare et al. (1989) list 5 diseases of T. billardierii caused by parasites and other agents.
HUSBANDRY
Thylogale billardierii is somewhat social, and 6– 10 adults of mixed sexes can be safely reared in a 0.5-ha enclosure. Fencing can be of wood or wire but must be at least 1.6 m high to prevent escape. In captivity, T. billardierii needs little shelter other than a roof, but large dog kennels and shrubbery are ideal in a large enclosure. Bulk feeders that release oats, specialized Kangaroo pellets (Wombaroo®, Wombaroo Food Products, South Australia, Australia), or similar pellets used to feed cattle provide most of the diet when supplemented every few days with fresh vegetables. Water, especially important when the major food source is dry, can be provided in regularly cleaned buckets or basins with dripping faucets.
Capture of captive T. billardierii is by use of a large butterfly net with strong netting. Animals tend to run along fence lines, and the net is suddenly pushed toward the fence as an animal runs by. This method is effective and reduces the stress of extended periods of chasing, which should be avoided. If collapse due to sudden fatigue does happen, rapid recovery can be achieved by an intramuscular injection of 10% glucose solution. An animal is easily handled when held firmly by the tail before placing it into a burlap bag and then using the handler’s knees to control the animal once on the ground. Pouch young can be safely removed from the teat from weeks 5 to 6 for the purpose of age estimation and weighing; its mother will usually accept the return of her young afterwards. While the young is temporarily out of the pouch, milk for analysis can be collected.
BEHAVIOR
Primarily nocturnal, Thylogale billardierii is sometimes seen during the day, especially toward dusk. It often signals the start of a period of foraging by drinking ( Morton and Burton 1973).
In locomotion, T. billardierii holds its forelimbs close to the chest and with the body more parallel to the ground (and less erect) than larger macropodids. Movement is swift and with great agility ( Morton and Burton 1973). The tail is not used for support when grazing, as happens in many macropodids.
Two visual signals were noted by Clancy (1982): grasspulling and standing full-stretch. The former behavior involves the assumption of erect posture, followed by grasping clumps of grass with the forepaws. Then the animal assumes a standing full-stretch posture, while tossing the grass tufts against the chest. This behavior often is repeated several times ( Clancy 1982). Clancy recorded 20 different behaviors, most usually initiated by the dominant animal.
Growling and clucking are the primary vocalizations. The former is a low-intensity sound made by the dominant animal in an aggressive interaction, and the latter is made in a range of situations, perhaps clouding its true role in communication ( Morton and Burton 1973). Three vocalizations recorded as sonograms by Clancy (1982) were hiss-growls, uttered by both sexes after physical contact; chucks, given by a male pursuing a female or supplanting another male and by females when signaling their young; and hisses, emitted by young when outside the pouch. T. billardierii also makes a nonvocal sound, foot thumping, when disturbed.
Aggressive interactions between adult males invariably lead to the smaller male retreating, whereas female–female interactions usually are won by the female with pouch young ( Morton and Burton 1973). The ritualized bouts between males resemble those of larger macropodids, including the boxing posture. Hair loss in males is especially common during the mating season, an indication that agonistic behavior is the likely cause.
A consequence of the dominance of the larger male, widespread in macropodids (including T. billardierii — Clancy 1982), is the winne-specific versus loser-specific behavior in prolonged bouts ( Ganslosser 1989). Fights normally are initiated by the prospective winner; the winner also tends to resume the fighting after a lull. Losers tend to end fights, to initiate lulls or breaks, and to use kicks earlier or more often than winners. Furthermore, losers tend to wrestle, attack, and embrace from lateral positions, winners from frontal positions. These ritualized fights seem important in maintaining or stabilizing dominance relationships, female fights being less ritualized ( Ganslosser 1989).
Males routinely exhibit sexual behavior by their daily olfactory inspections of females in their realm. Physical contact leads to courtship, escape by and pursuit of females, and attempted mountings, often 1 or 2/min for several minutes, before the male moves on. Females are receptive to being courted for a short time (<24 h; Rose and McCartney 1982a). If the female does not flee, the male follows her while emitting a clucking sound. Later, he sniffs the pouch and genital region.
During the often-lengthy copulation, the male holds the female with his forepaws around her flanks. Each copulation is characterized by a series of mountings, between which the male dismounts and rests for 4–15 min. This copulatory behavior is unusual for macropodids ( Morton and Burton 1973). However, one copulation observed by McCartney (1978) was different. Intromission at 5 min was followed by the male briefly retaining his grip on the female, after which she could move away. After 5 min spent licking the genital regions and autogrooming, each resumed grazing.
After the young leaves the pouch for the 1st time (6–7 months), it re-enters the pouch often for about 5 weeks, after which that privilege is suddenly withdrawn ( Morton and Burton 1973). Weaning, when it occurs 4 months later, is unpleasant for the youngster because the mother’s aggression increases quickly from cuffing to outright chases. Future mother–offspring interactions involve frequent mutual grooming, each bout consisting of cleaning the head and neck with the forepaws and tongue. In all, the lengthy period of development means that each female invests heavily in production of young.
Autogrooming, a prominent daytime activity, involves licking and scratching with forepaws and hind feet. The pouch and abdominal region are groomed by licking, often after hairs are parted with the forepaws. The hind feet scratch and groom the head and shoulders, often aided by the muscularity and flexibility of syndactylous digits 2 and 3. While at rest, T. billardierii often autogrooms for 1–10 min, and even more extensive grooming precedes the period of active foraging ( Morton and Burton 1973). Mutual allogrooming between mother and newly emerged young is one of the most common behaviors ( Clancy 1982). Clancy (1982) observed maternal aggression, a preliminary to weaning, as early as 288 days. In one instance, weaning occurred immediately.
GENETICS
The diploid number (2 n) of Thylogale billardierii is 22 chromosomes ( Sharman 1961). These include 2 large, 2 medium-sized, and 3 small acrocentrics, 1 metacentric, and 2 submetacentric pairs of autosomes, plus XX/XY sex chromosomes ( Rofe 1978).
CONSERVATION STATUS
Thylogale billardierii is among the critical weight range species that constitute the bulk of threatened and endangered Australian species that have disappeared or declined or are now restricted to islands ( Johnson et al. 1989). T. stigmatica and T. thetis , which are similar in size to T. billardierii , have stable populations on the mainland, and little is known about the population dynamics of the New Guinea Thylogale . T. billardierii is currently listed as “Least Concern” by the International Union for Conservation of Nature and Natural Resources ( Menkhorst and Denny 2016) and was considered secure in Tasmania until the intentional introduction of red foxes in 2001 or 2002. The presence of foxes probably contributed to the decline and disappearance of T. billardierii in Victoria and South Australia over a century ago and should the red fox become established in Tasmania, its threat to T. billardierii is real.
REMARKS
Thylogale is derived from the Greek words thylakos, meaning pouch, and gale, meaning weasel. The specific name billardierii is the Latin form of Dr. J. J. H. Labillardiere, a naturalist who accompanied the Bruni D’Entrecasteaux expedition to Van Diemen’s Land (later to be renamed Tasmania). He collected the 1st specimen there in 1792 ( Strahan 1981).
Other common names include rufous wallaby, red-bellied wallaby, red-bellied pademelon, rufous-bellied pademelon, scrub wallaby, and, in Tasmania, simply wallaby.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Thylogale billardierii ( Desmarest, 1822 )
| Rose, Randolph W. & Rose, Robert K. 2018 |
Halmaturus (Thylogale) tasmanei
| Gray 1838: 108 |
Macropus (Halmaturus) rufiventer
| Ogilby 1838: 220 |