
Rayenia malalcurensis Menegoz
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.484.1.4 |
|
persistent identifier |
https://treatment.plazi.org/id/9746F54B-511D-B149-9FFC-FD64806EFEB1 |
|
treatment provided by |
Marcus |
|
scientific name |
Rayenia malalcurensis Menegoz |
| status |
gen. nov., sp. nov. |
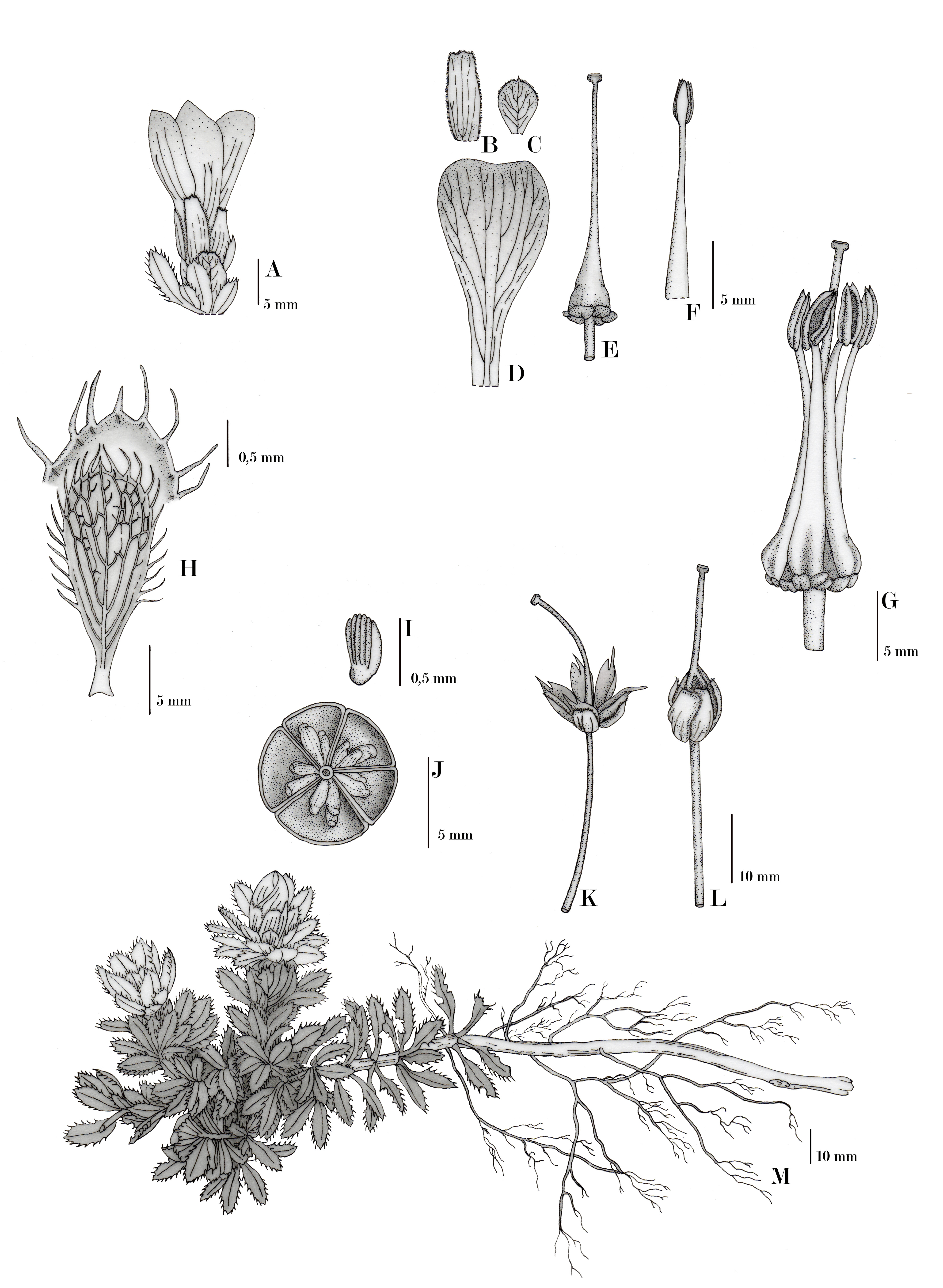
Rayenia malalcurensis Menegoz View in CoL & A.E. Villarroel, gen. nov., sp. nov. ( Figures 3–4 View FIGURE 3 View FIGURE 4 ).
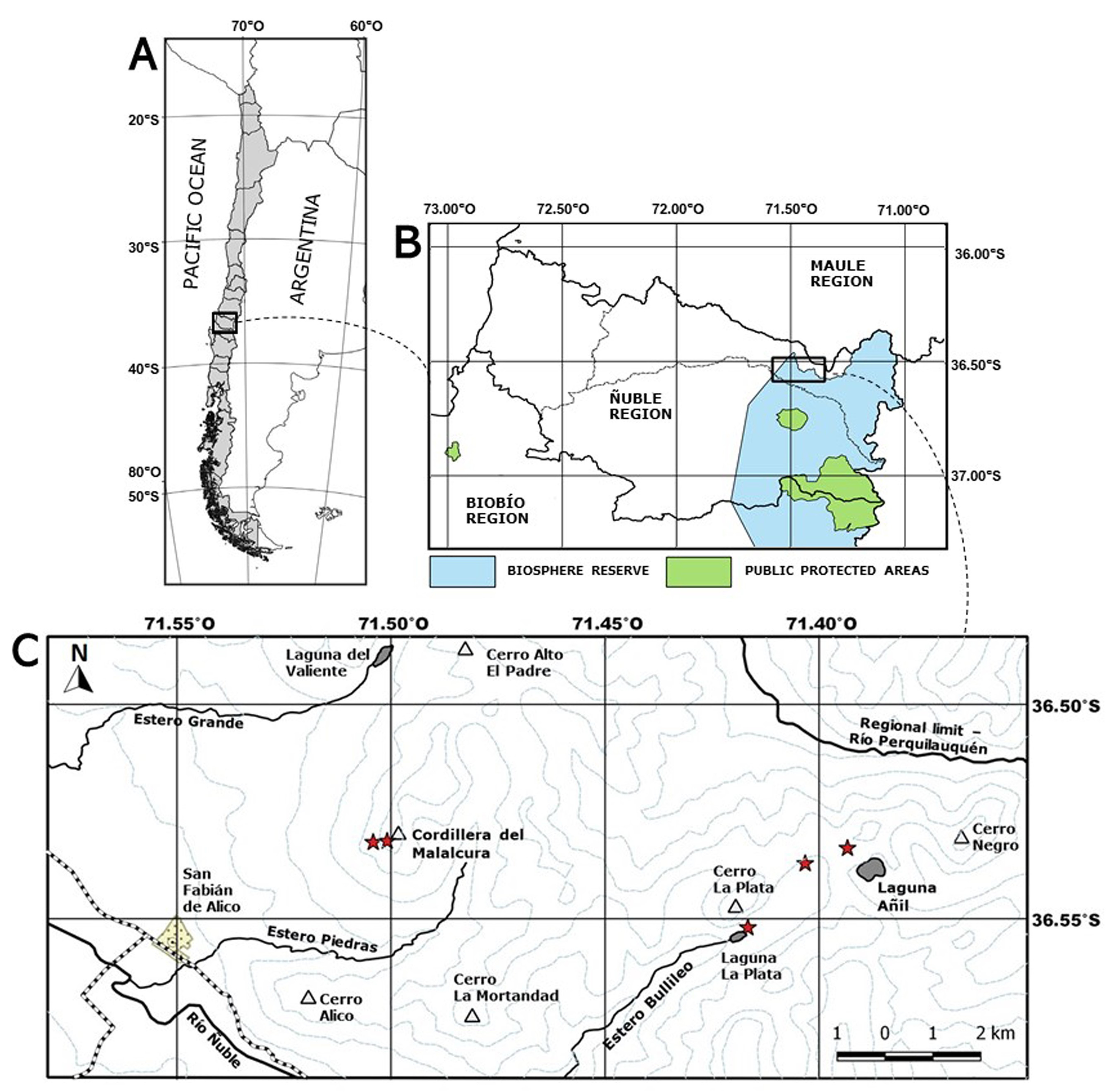
Type: — CHILE. Ñuble Region , Punilla Province , San Fabián de Alico, Cordillera del Malalcura, crevices of granite rocks, 1747 m elevation, 36°31’56.03” S, 71°30’14.02” W, 11 October 2018, K GoogleMaps . Menegoz , G . Ossa & J. C . Covarrubias 121, (holotype SGO!; isotypes CONC!, ULS!, VALD!, E!) .
Diagnosis: — Rayenia malalcurensis is similar to Tribeles australis , but differs by its leaf apex that is mucronate (vs. tridentate), leaf margin finely serrate and ciliated (vs. entire), sepals free (vs. basally united), petals pink (vs. white), anthers adnate (vs. basifixed), ovary 5-carpelate, pentalocular (vs. 3-carpelate, trilocular), stigma discoid (vs. trilobed), capsule septicidal (vs. loculicidal), entire sepals and style marcescent (vs. remains of sepals and style marcescent), and seeds pale mustard yellow and expelled from the locules of mature fruits (vs. black and remaining attached to central fruit axis).
Description: —Perennial creeping subshrub, usually forming cushions of variable size, sometimes with unclear limits between neighbouring individuals, 5–20 cm tall, 20–180 cm wide, 50–1050 cm long. Roots extended, woody and ramified; taproot brown, circular, 1–2.2 mm diameter, with small secondary roots, brown, 0.1–0.5 mm diameter. Main branches ligneous, prostrate, circular, up to 5 cm diameter; bark grey to light brown, corrugated and stringy texture; secondary branches usually prostrate, brown, circular, 1–4.5 mm diameter, radially arising and dichotomously branching; terminal shoots erect, with 14–46 terminal leaves, under which some dry leaves persist. Leaves lustrous green, leaf base white, entire, sessile, and arranged in an imbricate rosette. Lamina oblanceolate to scarcely obovate, flat, 6.3–19.9 × 2.2–7.4 mm, gradually reducing in size towards the centre of the rosette; base decurrent; apex mucronate, sometimes ciliate; mucro white turning to yellow-brown, 0.5–0.8 mm long; margin finely serrate towards the upper 2/3 of the blade, with up to 15 teeth on each side, usually ciliated; cilia white turning to yellow-brown, 1–1.8 mm long; coriaceous, glabrous on both sides; venation non prominent, pinnate, festooned semicraspedodromous. Inflorescence a terminal solitary flower, surrounded by bracts and the upper rosette leaves, some of them in transition from leaves to foliaceous bracts. Foliaceous bracts 2–4, light green, slightly darker towards the apex, ovate, 2.5–8 × 6– 1.3 mm, base white, truncate and decurrent, apex cuspidate, margin minutely pubescent, texture membranous. Flowers terminal, pink, actinomorphic, campanulate, contort, dichlamydeous, hermaphrodite, 2.8–3 × 2.4 mm. Pedicels green, circular and short, 0.2–0.3 mm long. Receptacle green towards the centre and pink on the outer edge, discoid, flat to slightly convex, 0.4–0.6 mm diameter. Sepals 5, free, variable in colour, from rose changing to cream or yellow-green, oblongate, 3.6–6.5 × 9.5–13 mm, slightly wider towards the apex, base truncate, apex truncate and membranous, margin irregularly dentate, minutely pubescent, texture membranous, marcescent on fruit. Petals 5, free, imbricate, pale pink with darker pink base and veins, oblanceolate, slightly concave, 10.6–16 × 24.2–31 mm, base cuneate, apex rounded to truncate, margin entire, membranous texture. Stamens 5, free, haplostemonous, filaments white and pink at base, linear and widest at base, 17.1–19 mm long. Anthers yellow, oblong, 3.4–4 mm long, adnate, extrorse, longitudinally dehiscent. Ovary superior, green, darker at its base, conical, 0.5 × 0.8 mm, glabrous, syncarpous. Carpels 5. Locules 5. Placentation axile, placentae 2 per locule. Style 1, variable in colour, from white changing to pale pink or green, linear, 16.9–20.6 mm long, marcescent on fruit; stigma discoid green to brown. Fruit a septicidal 5-valved capsule, yellow-green colour when immature, ovoid with terminal persistent style, 6–10 × 11.3–15.3 mm (without style), glabrous, surrounded by persistent sepals. Mature fruit (open and dry), brown, star-shaped, ligneous texture, with persistent dry style and sepals, some remaining on the plant from one year to the next; pedicels usually grow during the fruit maturation process, reaching 38–53 mm long. Seeds small, 11–24 per valve, pale mustard yellow, with varying shapes but mainly oblong, circular to elliptical in cross-section, 0.5 × 1.5–2 mm, surface ribbed.
Etymology: —We named the genus and specific epithet after the Mapudungun (Mapuche language) to honour the indigenous Mapuche people, who live in central-southern Chile and Argentina, where traditionally they have very close spiritual links to nature. The genus Rayenia comes from “rayen” which means “flower”, to highlight the remarkable flower of this species. While the epithet malalcurensis derives from “Malalcura’’ that means “rock enclosure”, referring to the type specimen locality (Cordillera del Malalcura), which is a hill crowned by granite rocks. Altogether, Rayenia malalcurensis means “flower of rock enclosure”, from the spanish “flor de corral de piedra”.
Phenology: —Flowering from end of September to November; fruiting from November to January.
Distribution and habitat: —Endemic to the eastern Andean ranges in the locality of San Fabián de Alico, Punilla Province, Ñuble Region, Chile ( Fig. 1C View FIGURE 1 ). Specifically, 5 sites are known: two of them located in the Cordillera del Malalcura (1887 m elevation, 36°31’56” S 71°30’14” W; 1722 m elevation, 36°31’55” S 71°30’14” W), two others in the Valle del estero Bullileo (1270 m elevation, 36°33’7” S 71°24’59” W; 1413 m elevation, 36°32’13” S 71°24’11” W); and the last one near Laguna Añil (1733 m elevation, 36°32’ S 71°23’36” W). The maximum distance between the sites is 10 km.
The species usually grows at high elevations (1270–1900 m), in crevices and small terraces of south-southwest facing granite rocks, with variable slopes (0° to 80°) ( Fig. 5 View FIGURE 5 ). Due to snow melting and low sun exposure, these sites remain humid during the dry season. Few plants were observed growing in the shade of neighbouring shrubs or trees. Two weather stations, Punilla (840 m elevation) and Caracol (725 m elevation), indicate that the average annual temperature and precipitation is 11.8 °C and 2150 mm, respectively, for the period 1965–2012 ( DGA 2018). In addition, a private pluviometer in the village of San Fabián de Alico (447 m elevation) recorded an annual average precipitation of 1434 mm for the period 2017–2019 (Covarrubias J.C., pers. comm.).
Associated vegetation: —From the phytogeographic point of view, Rayenia malalcurensis is part of the vegetal formation known as Andean deciduous forest of the Bío-Bío ( Gajardo 1994). Another classification considers that there are two vegetative belts in this area ( Luebert & Pliscoff 2017): (1) Andean mediterranean deciduous forest of Nothofagus obliqua ( Mirbel 1827: 465) Ørsted (1871: 354) (Nothofagaceae) and Austrocedrus chilensis (D.Don in Lambert 1832: 128) Pichi Sermolli & Bizzarri (1978: 482) ( Cupressaceae ); and (2) Andean temperate deciduous forest of Nothofagus pumilio ( Poeppig & Endlicher 1838: 68) Krasser (1896: 161) and Azara alpina Poeppig & Endlicher (1838: 30) (Salicaceae) .
Our field observations indicate a total of 70 species associated with Rayenia malalcurensis . In some areas as in Cordillera del Malalcura (1887 m elevation) R. malalcurensis can be associated with only 4 species, and the richest site is in the Valle del estero Bullileo (1413 m elevation) with 33 species. The most abundant species are Chusquea montana Philippi (1864: 298) (Poaceae) , Myrteola nummularia (Poir in Lamarck 1798: 407) Berg (1856: 396) ( Myrtaceae ), Luzula spp . de Candolle (1805: 158) ( Juncaceae ), N. pumilio , Festuca spp . Linnaeus (1753: 73) ( Poaceae ), Orites myrtoidea ( Poeppig & Endlicher 1838: 35) Bentham & Hooker ex Engler (1889: 146) (Proteaceae) , N. obliqua , Eucryphia glutinosa ( Poeppig & Endlicher 1838: 68) Baillon (1869: 461) (Cunoniaceae) , Gaultheria poeppigii de Candolle (1839: 593) (Ericaceae) , Empetrum rubrum Vahl ex Willdenow (1806: 713) (Ericaceae) , Carex spp . Linnaeus (1753: 972) ( Cyperaceae ), Haplopappus spp . Cassini (1828: 56) ( Asteraceae ) and Gaultheria tenuifolia (Philippi 1858: 10) Sleumer (1936: 207) .
Additional specimens examined (paratypes): — CHILE. Ñuble Region , Punilla Province , San Fabián de Alico : Valle del estero Bullileo – Laguna La Plata, 13 February 2019, A. E . Villarroel, R . Neira & E . Ponce ( SGO!), 10 November 2019, A . Villarroel, D. Quinchero & C . Lafquén ( EIF!), 10 January 2020, A. E . Villarroel & A . Villarroel A. ( VALD!); Valle del estero Bullileo, 9 November 2019, A. E . Villarroel, S . Castro & D. Acevedo ( SGO!), 8 January 2020, A. E . Villarroel & E . Ponce ( CONC!); in the vicinity of Laguna Añil , 7 January 2020, A. E . Villarroel & E . Ponce ( ULS!) .
Conservation status: — Rayenia malalcurensis is assessed here as Critically Endangered (CR) under the IUCN categories and criteria B1ab(iii); C2a(ii). Criterion B1 was selected because its extent of occurrence is <100 km 2 (10.634 km 2). Criterion “a” was selected because it is known to exist in only a single location, and a single population with 5 sites located no further than 10 km away from each other. Criterion “b(iii)” was selected because there is a projected decline in the area, extent, and quality of habitat. Criterion C2(a)(ii) was selected because there are less than 250 mature individuals, all of which occur in a single population. So far, 51 individuals have been recorded at 5 sites, ranging from 1 to 34 individuals per site. High Andean plants are very sensitive to global warming, given that migration is limited by the lack of connection between summits. There has been a significant reduction in precipitation and snow cover, together with an increase in temperatures in Central Chile for the last decade ( Garreaud et al. 2017, Cordero et al. 2019), and climate change scenarios project an increase of at least 1° C of the mean temperature for the next 30 years, plus a decrease in winter precipitations of about 30% at the end of the century ( Bozkurt et al. 2017). Since the species occurs near the summits of a mountain range with very specific soil types and exposition, we project a continuing decline in the area and quality of habitat in the coming decades. Rayenia malalcurensis is not present in any protected area in Chile ( Fig. 1B View FIGURE 1 ).


| K |
Royal Botanic Gardens |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| J |
University of the Witwatersrand |
| C |
University of Copenhagen |
| SGO |
Museo Nacional de Historia Natural |
| CONC |
Universidad de Concepción |
| ULS |
Universidad de La Serena |
| VALD |
Universidad Austral de Chile |
| E |
Royal Botanic Garden Edinburgh |
| A |
Harvard University - Arnold Arboretum |
| R |
Departamento de Geologia, Universidad de Chile |
| EIF |
Universidad de Chile |
| S |
Department of Botany, Swedish Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |